|
|
Aconitasi e |
|||||
|
Molecola del Mese di Maggio 2007 L'aconitasi catalizza la seconda reazione del ciclo di Krebs, ma, nel citoplasma è una proteina regolatrice dei livelli del ferro 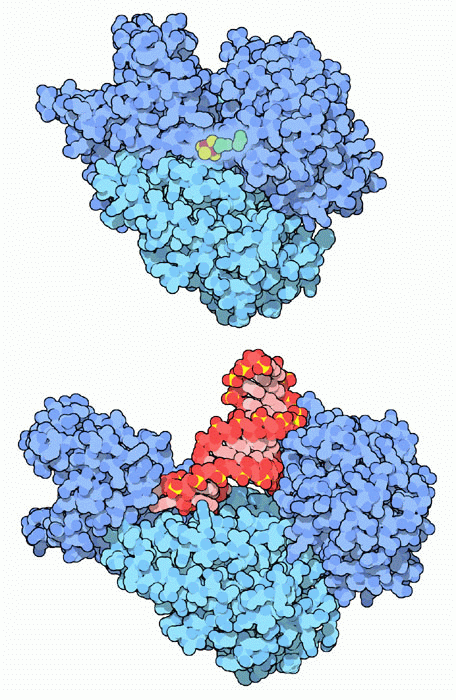 Introduzione IntroduzioneL'aconitasi è un enzima essenziale nel ciclo di Krebs (mdm 10-2012) e la proteina 1 regolatrice del ferro interagisce con l'RNA messaggero per controllare i livelli di ferro all'interno delle cellule. Ci si potrebbe chiedere: cos'hanno in comune queste due proteine? Sono state scoperte da ricercatori diversi che hanno assegnato loro nomi che descrivevano le loro due funzioni del tutto diverse. Sorprendentemente, però, quando hanno determinato la sequenza di amminoacidi di queste due proteine, hanno scoperto che erano identiche. Quindi la stessa proteina svolge due compiti del tutto diversi! Aconitasi . . . L'enzima aconitasi ha un ruolo chiave nella respirazione cellulare, la via principale di produzione dell'energia. E' uno degli enzimi del ciclo di Krebs e converte l'acido citrico in acido isocitrico. Il ciclo di Krebs è localizzato nei mitocondri, è qui che l'enzima aconitasi trasforma l'acido citrico in isocitrico, ma esiste un'altra forma simile di aconitasi che opera nel citoplasma e che genera acido isocitrico per altri scopi sintetici. La forma citoplasmatica dell'aconitasi è mostrata qui a destra in alto (file PDB 2b3y). E' composta da una singola catena proteica che si avvolge formando più domini (azzurro e blu). I domini si stringono come una morsa attorno al sito attivo, che contiene un complesso ferro-zolfo per catalizzare la reazione. . . . o proteina 1 regolatrice del ferro La forma citoplasmatica dell'aconitasi agisce anche come proteina 1 regolatrice del ferro. La struttura qui a destra (file PDB 2ipy) ci mostra come viene realizzata questa azione del tutto diversa. Il complesso ferro-zolfo dell'aconitasi è legato in modo instabile e deve essere sostituito di tanto in tanto. Quando i livelli di ferro nella cellula diventano troppo bassi, non c'è abbastanza ferro per rigenerare il complesso ferro-zolfo e la proteina passa alla sua seconda funzione. La proteina assume una forma più aperta e si lega a specifici tratti a forcina di alcuni particolari mRNA. Viene legato una tratto a forcina all'inizio dell'RNA messaggero che codifica per la ferritina e cinque tratti a forcina alla fine dell'RNA messaggero che codifica per il recettore della transferrina. In questa forma l'enzima è diventato la proteina 1 regolatrice del ferro e inibisce la formazione di ferritina, così meno ferro viene tenuto fermo in deposito, ed aumenta la sintesi del recettore della transferrina, così la cellula può catturare dal sangue più transferrina e, con essa, più ferro. Nell'immagine qui sotto, realizzata con Chimera, si vedono quattro basi azotate dell'mRNA ruotate in fuori e legate nel sito attivo dalla proteina 1 (file PDB 3snp). 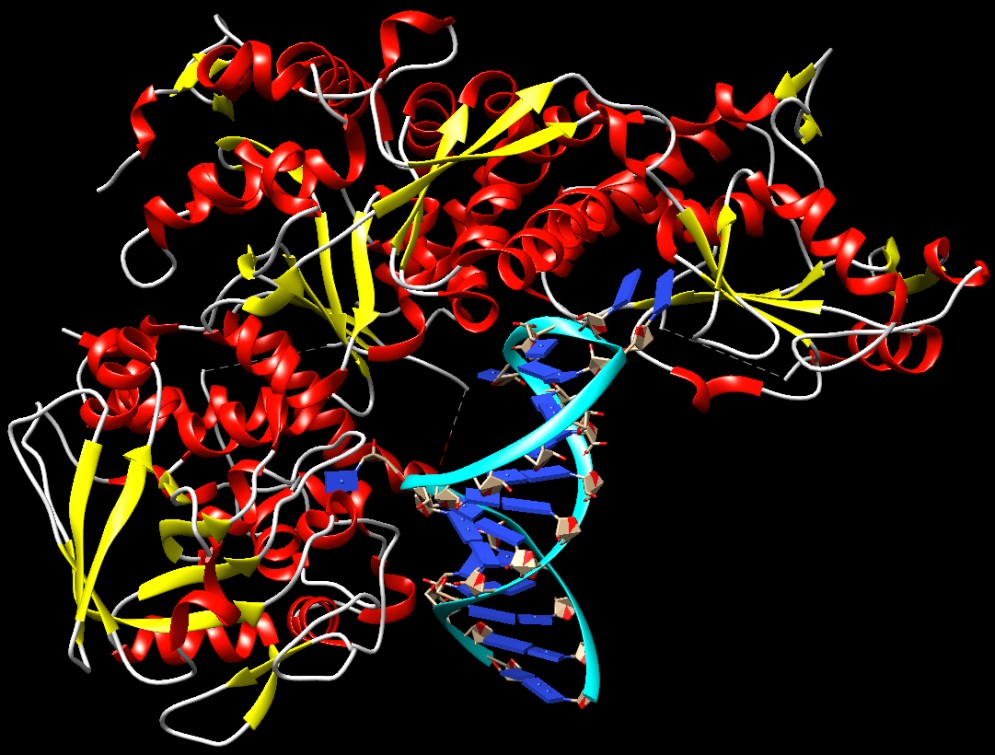
Proteine dalla doppia personalità 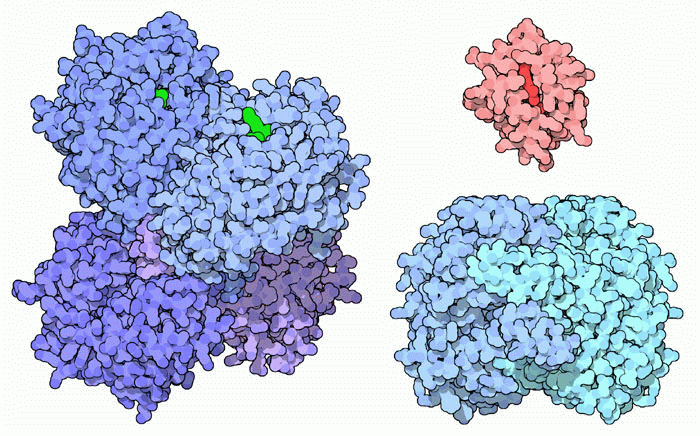 Molte
altre proteine hanno una doppia vita e svolgono due funzioni del tutto
diverse tra loro. Qui ne mostriamo tre esempi. Molte
altre proteine hanno una doppia vita e svolgono due funzioni del tutto
diverse tra loro. Qui ne mostriamo tre esempi. L'enzima retinale deidrogenasi che converte il retinale in acido retinoico, una molecola regolatoria, è mostrato qui a lato sulla sinistra (file PDB 1o9j) con il coenzima NADH in verde. La sua seconda funzione è quella di modificare la consistenza e la trasparenza del cristallino, la lente dell'occhio. Il citocromo c (mdm 12-2002) è mostrato in alto a destra (rosso) (file PDB 3cyt). Svolge un ruolo chiave nella produzione di energia, trasportando elettroni nella catena di trasporto di elettroni nei mitocondri. Quando però la cellula è danneggiata, il citocromo c esce dal mitocondrio e va nel citoplasma dove svolge la sua seconda funzione: dà inizio ad una cascata di eventi che alla fine porta alla morte programmata della cellula, l'apoptosi. La glucosiofosfato isomerasi (file PDB 2pgi), la terza proteina dalla doppia personalità (azzurra), è uno dei dieci enzimi che realizzano la glicolisi (mdm 1-2004). Questa proteina può essere anche secreta all'esterno della cellula dove agisce come messaggero cellulare assumendo diversi nomi: neuroleukina, fattore di mobilità autocrina, e mediatore di differenziazione e maturazione. Questi nomi ci possono dare un'idea dei diversi messaggi che questa proteina può portare fuori dalla cellula. 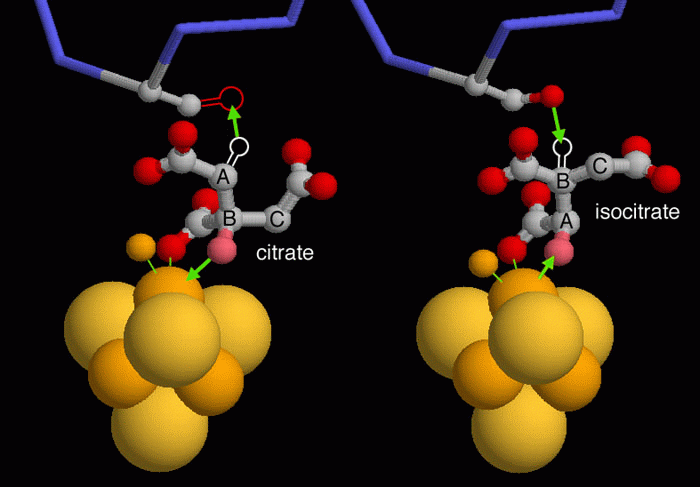 Esplorando
la struttura Esplorando
la strutturaL'aconitasi realizza una classica reazione stereospecifica che viene spesso usata come esempio nei libri di biochimica. Estrae un gruppo idrossilico (OH) ed uno specifico atomo di idrogeno dall'acido citrico e li inserisce in una nuova posizione geometricamente determinata per formare l'acido isocitrico. Questo processo è illustrato nelle due strutture cristalline qui a fianco, ma è necessaria un po' di immaginazione per interpretarle correttamente perchè le strutture non mostrano gli atomi di idrogeno. La molecola mostrata sulla sinistra, (file PDB 1c96) ha l'acido citrico legato nel sito attivo. Nella forma normale dell'enzima, l'atomo di ossigeno rosa sul carbonio B, viene estratto verso il basso dal complesso ferro-zolfo, mentre un atomo di idrogeno dovrebbe essere estratto verso l'alto dall'ossigeno di una serina (queste due reazioni sono evidenziate con frecce verdi). In questa struttura, però la serina è stata sostituita con una alanina che non possiede l'atomo di ossigeno necessario per l'estrazione dell'idrogeno (è proprio grazie a questa mutazione che il sito attivo si è potuto congelare in questa posizione per ottenerne un cristallo: la reazione non può continuare senza l'ossigeno della serina!). La seconda fase della reazione è illustrata qui sopra sulla destra (file PDB 7acn). La molecola di acido citrico disidratato si è capovolta nel sito attivo (notate la differente posizione delle lettere A, B, C). L'atomo di ossigeno è rientrato da sotto sul carbonio A mentre l'atomo di idrogeno è rientrato da sopra sul carbonio B, quindi si trovano in posizioni invertite rispetto a prima e hanno formato acido isocitrico. P. J. Artymiuk, J. Green (2006) The double life of aconitase. Structure 14, 2-4. T. A. Rouault (2006) The role of iron regulatory proteins in mammalian iron homeostasis and disease. Nature Chemical Biology 2, 406-414. C. J. Jeffery (2004) Molecular mechanisms of multitasking: recent crystal structures of moonlighting proteins. Current Opinion in Structural Biology 14, 663-668. S. D. Copley (2003) Enzymes with extra talents: moonlighting functions and catalytic promiscuity. Current Opinion in Chemical Biology 7, 265-272.
|
||||||
|
|
||||||
