|
|
Ciclo dell'acido citrico |
|||||
|
Molecola del Mese di Ottobre 2012 Il ciclo dell'acido citrico o ciclo di Krebs è costituito da 8 tappe enzimatiche ed ha un ruolo essenziale nella produzione di energia 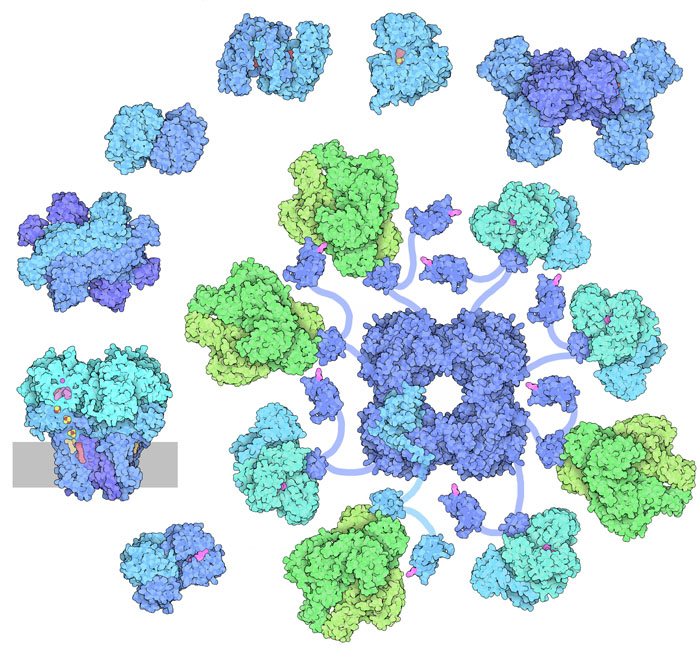 Introduzione IntroduzioneIl ciclo dell'acido citrico, conosciuto anche come ciclo di Krebs o ciclo degli acidi tricarbossilici, ha un ruolo centrale nel metabolismo della cellula sia nella produzione di energia che per la sintesi di nuove molecole. Il ciclo di Krebs costituisce la terza e ultima tappa delle reazioni di demolizione del glucosio, dopo la glicolisi (mdm 2-2004) e la decarbossilazione ossidativa (mdm 9-2012). Inoltre fornisce NADH e FADH2 alla catena respiratoria (mdm 12-2011) e alla fosforilazione ossidativa (mdm 12-2005) che, facendoli reagire con ossigeno, producono grandi quantità di ATP. Il ciclo dell'acido citrico è anche importante nella biosintesi, perchè fornisce intermedi per la sintesi di amminoacidi e di altre molecole. Gli enzimi del ciclo dell'acido citrico si trovano in tutte le cellule che usano ossigeno O2 e persino in alcune cellule che non lo usano. Gli enzimi mostrati in questo articolo sono stati isolati da molti organismi diversi. Nella figura qui a fianco si vedono gli otto enzimi uno vicino all'altro. Appare evidente che uno degli enzimi ha dimensioni enormi rispetto agli altri, ma in realtà è ancora più massiccio di così perchè la subunità centrale è circondata da ben 24 subunità, mentre qui ne sono mostrate solo 8. Otto reazioni Le otto reazioni del ciclo dell'acido citrico hanno lo scopo di trasformare l'acetil Coenzima-A in CO2 e completare così l'ossidazione del glucosio, cominciata con la glicolisi e la decarbossilazione ossidativa. Per questo viene usato, come catalizzatore, l'ossalacetato, un acido bicarbossilico che contiene anche un gruppo chetonico. Il ciclo comincia legando il gruppo acetile all'ossalacetato, si forma così acido citrico, un acido tricarbossilico che dà il nome a tutto il ciclo. Poi, in otto tappe, il gruppo acetile viene completamente ossidato e trasformato in due molecole di CO2 mentre la parte restante della molecola viene riconvertita in ossalacetato per cominciare un nuovo ciclo. Queste otto reazioni, però, non sono semplici come sembrano. Ci si potrebbe aspettare che gli enzimi eliminassero i due carboni del gruppo acetile usando l'ossalacetato solo come supporto. Invece, marcando alcuni atomi di carbonio nelle due molecole, gli scienziati hanno scoperto che le cose vanno in modo diverso. Infatti i due atomi che vengono persi come CO2 appartengono all'ossalacetato, mentre il gruppo acetile va a costituire la struttura del nuovo ossalacetato che si forma. Centrale energetica Il ciclo dell'acido citrico fornisce gli elettroni (sotto forma di NADH) necessari al processo della fosforilazione ossidativa che produce la maggior parte dell'energia nelle nostre cellule sotto forma di ATP. Quando il gruppo acetile viene ossidato a CO2, cede elettroni al NADH e questo a sua volta li cede al Complesso 1 (mdm 12-2011) della catena respiratoria. Questi elettroni da qui giungono all'ossigeno O2 attraverso una serie di tre pompe protoniche situate, la prima, a ridosso del complesso 1, la seconda, nel complesso 3 (citocromo bc1 (mdm 5-2011)) e, la terza, nel complesso 4 (citocromo c ossidasi (mdm 12-2002)). Le pompe protoniche generano un gradiente di ioni H+ a cavallo della membrana interna dei mitocondri che mette in rotazione l'enzima ATP sintasi (mdm 12-2005) che è la macchina proteica che sintetizza ATP. Tutti i complessi della catena respiratoria e l'enzima ATP sintasi si trovano immersi nella membrana interna dei mitocondri. 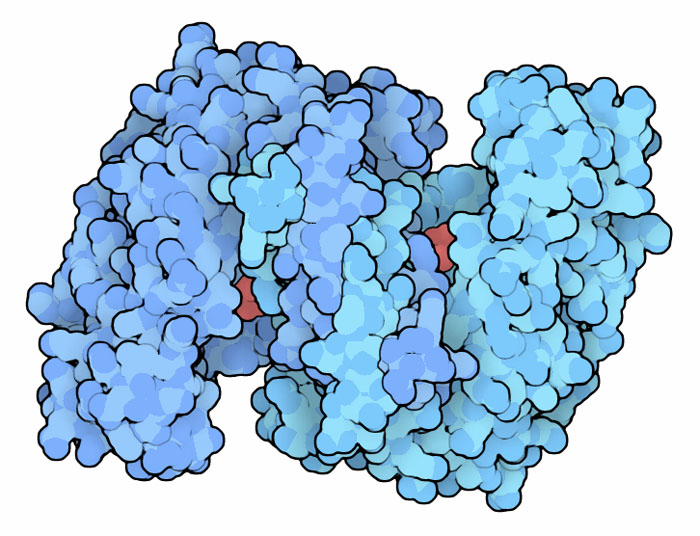 (1) Citrato sintasi Il ciclo di Krebs, o ciclo dell'acido citrico, inizia con l'enzima citrato sintasi, mostrato qui a destra (file PDB 1cts). Questo enzima prende l'acetil coenzima A (generato dalla decarbossilazione ossidativa dell'acido piruvico) e lo lega all'ossalacetato formando acido citrico. L'enzima si apre e si chiude attorno a queste molecole durante la reazione. Per vederlo in azione andate alla molecola del mese 9-2007 (citrato sintasi). 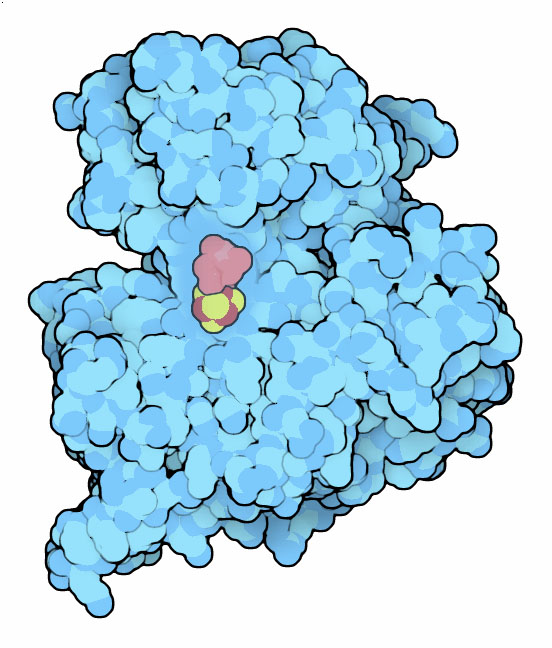 (2) Aconitasi L'acido citrico formato nel primo stadio non è facilmente decarbossilabile perché ha il gruppo alcolico al centro della molecola, in posizione terziaria, dove non può essere ossidato. Per questo nel secondo stadio, il gruppo OH viene spostato sul carbonio in fianco, in posizione secondaria. L'enzima aconitasi, mostrato qui a fianco (file PDB 7acn), realizza la reazione di isomerizzazione con l'ausilio di alcuni gruppi ferro-zolfo. Per vedere l'enzima in azione andate alla molecola del mese 5-2007 (aconitasi). (3) Isocitrato deidrogenasi 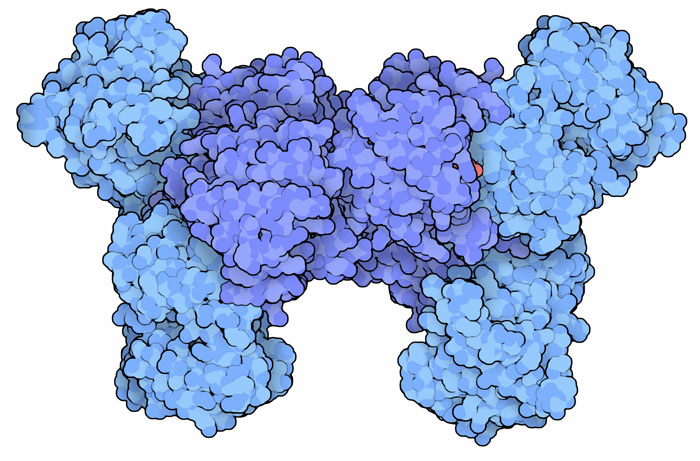 Il
vero lavoro di decarbossilazione comincia nella terza tappa del ciclo.
L'enzima isocitrato deidrogenasi, mostrato qui a fianco (file PDB 3blw)
prima ossida l'acido isocitrico trasformando l'OH della molecola in carbonile
e poi rimuove il carbossile centrale dell'acido. Durante l'ossidazione
trasferisce gli elettroni al NADH. Per vedere l'enzima in maggior dettaglio,
andate alla molecola del mese 9-2010 (isocitrato
deidrogenasi). Il
vero lavoro di decarbossilazione comincia nella terza tappa del ciclo.
L'enzima isocitrato deidrogenasi, mostrato qui a fianco (file PDB 3blw)
prima ossida l'acido isocitrico trasformando l'OH della molecola in carbonile
e poi rimuove il carbossile centrale dell'acido. Durante l'ossidazione
trasferisce gli elettroni al NADH. Per vedere l'enzima in maggior dettaglio,
andate alla molecola del mese 9-2010 (isocitrato
deidrogenasi).(4) Complesso 2-oxoglutarato deidrogenasi 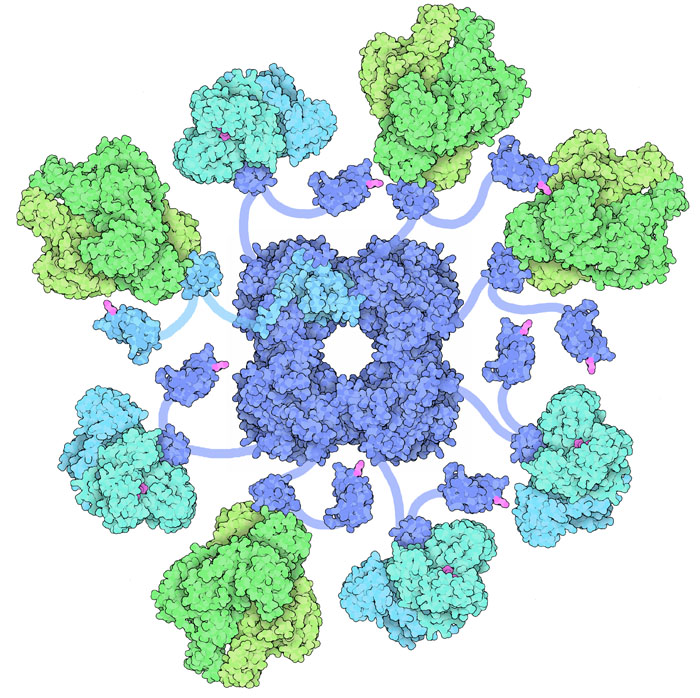 Il
quarto stadio è realizzato da un grande complesso multienzimatico,
simile al complesso della piruvato
deidrogenasi (l'enzima della decarbossilazione ossidativa dell'acido
piruvico, mdm 9-2012). Infatti questo enzima realizza una reazione molto
simile alla classica decarbossilazione ossidativa e utilizza lo stesso
set di molecole: vitamina B1, acido lipoico, coenzima A, NADH. Nell'enzima
si realizzano più reazioni in sequenza. Dapprima avviene una decarbossilazione
con l'ausilio della vitamina B1 (tiaminapirofosfato), poi avviene l'ossidazione
da parte dell'acido lipoico, poi il carbossile così ottenuto si
lega al coenzima A, infine l'acido lipoico ridotto trasferisce elettroni
al NADH. Il complesso è formato da tre enzimi diversi connessi
da braccia flessibili. L'illustrazione qui a fianco mostra solo alcuni
di questi enzimi legati al guinzaglio, ma nel complesso reale il corpo
centrale è circondato da 24 enzimi e ha l'aspetto di una grossa
palla multienzimatica. Questa figura è stata ottenuta usando molte
strutture PDB: 1e2o,
1bbl, 1pmr,
2eq7, e 2jgd. Per comprendere
meglio il funzionamento di questo grande complesso multienzimatico andate
alla molecola del mese 9-2012 dove è illustrato un enzima simile
piruvato
deidrogenasi. Il
quarto stadio è realizzato da un grande complesso multienzimatico,
simile al complesso della piruvato
deidrogenasi (l'enzima della decarbossilazione ossidativa dell'acido
piruvico, mdm 9-2012). Infatti questo enzima realizza una reazione molto
simile alla classica decarbossilazione ossidativa e utilizza lo stesso
set di molecole: vitamina B1, acido lipoico, coenzima A, NADH. Nell'enzima
si realizzano più reazioni in sequenza. Dapprima avviene una decarbossilazione
con l'ausilio della vitamina B1 (tiaminapirofosfato), poi avviene l'ossidazione
da parte dell'acido lipoico, poi il carbossile così ottenuto si
lega al coenzima A, infine l'acido lipoico ridotto trasferisce elettroni
al NADH. Il complesso è formato da tre enzimi diversi connessi
da braccia flessibili. L'illustrazione qui a fianco mostra solo alcuni
di questi enzimi legati al guinzaglio, ma nel complesso reale il corpo
centrale è circondato da 24 enzimi e ha l'aspetto di una grossa
palla multienzimatica. Questa figura è stata ottenuta usando molte
strutture PDB: 1e2o,
1bbl, 1pmr,
2eq7, e 2jgd. Per comprendere
meglio il funzionamento di questo grande complesso multienzimatico andate
alla molecola del mese 9-2012 dove è illustrato un enzima simile
piruvato
deidrogenasi.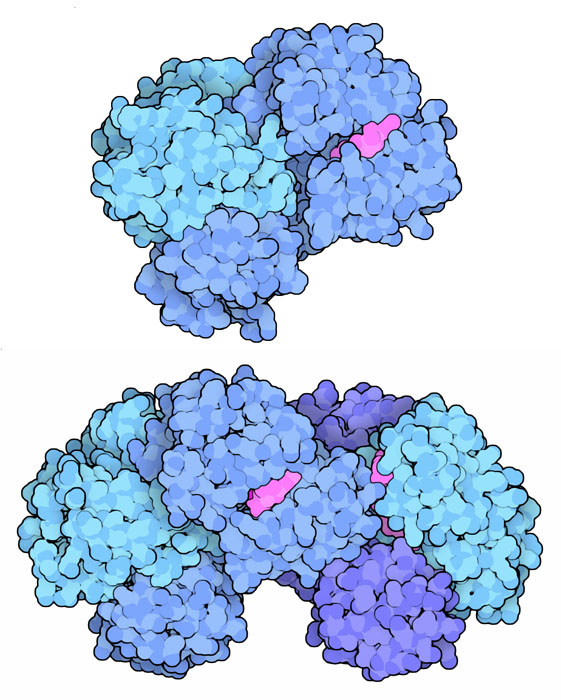 (5) Succinil-CoA sintetasi Il quinto stadio del ciclo di Krebs è l'unico nel quale viene prodotto ATP. Il legame tioestere tra succinato e coenzima A è molto instabile e, rompendosi, libera l'energia necessaria alla sintesi di ATP. In realtà, nei mitocondri, l'enzima (mostrato qui a destra in alto, file PDB 2fp4) produce GTP che viene poi convertito in ATP da un altro enzima, nucleotide difosfato chinasi. Nel citoplasma si trova una forma simile di succinil-CoA sintetasi che usa ATP (e non GTP) e che realizza la reazione inversa, sintetizza succinil-CoA che poi viene utilizzato in altre reazioni. Qui a fianco in basso è mostrata una forma batterica dell'enzima succinil-CoA sintetasi ATP-dipendente (file PDB 1cqi). (6) Succinato deidrogenasi 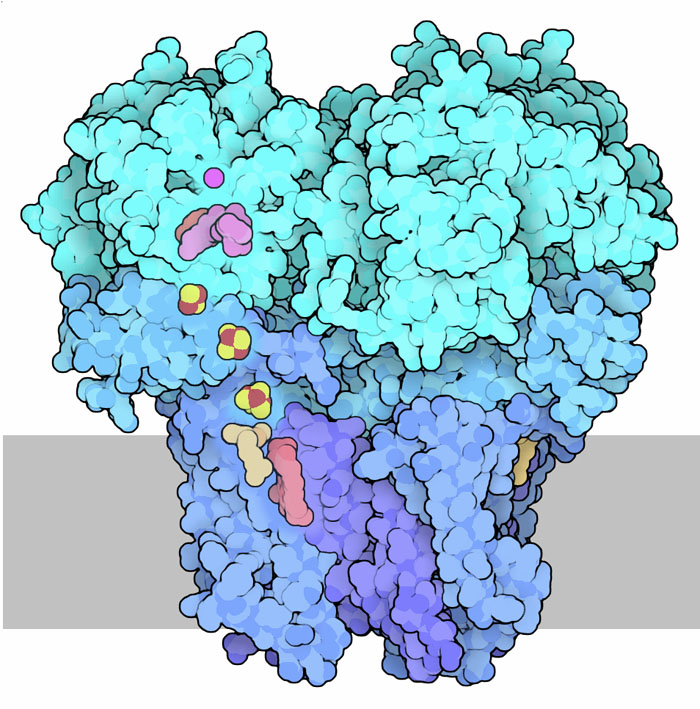 La
sesta tappa è realizzata da un complesso proteico legato alla membrana
interna dei mitocondri. Questo enzima realizza la sesta reazione del ciclo
di Krebs direttamente all'interno del complesso 2 della catena respiratoria.
L'enzima crea un doppio legame carbonio-carbonio nella molecola di acido
succinico ossidandolo ad acido fumarico e trasferisce al FAD (magenta
in alto nella figura) gli elettroni dei due atomi di idrogeno estratti.
Da qui gli elettroni vengono trasferiti attraverso una serie di centri
ferro-zolfo (gialli e rossi) e un gruppo eme (rosa) fino al coenzima Q
(beige chiaro), un trasportatore di elettroni mobile (dentro la membrana)
che porta gli elettroni al citocromo
bc1 (mdm 5-2011) nel complesso 3. L'enzima mostrato qui a destra
è batterico (file PDB 1nek). La
sesta tappa è realizzata da un complesso proteico legato alla membrana
interna dei mitocondri. Questo enzima realizza la sesta reazione del ciclo
di Krebs direttamente all'interno del complesso 2 della catena respiratoria.
L'enzima crea un doppio legame carbonio-carbonio nella molecola di acido
succinico ossidandolo ad acido fumarico e trasferisce al FAD (magenta
in alto nella figura) gli elettroni dei due atomi di idrogeno estratti.
Da qui gli elettroni vengono trasferiti attraverso una serie di centri
ferro-zolfo (gialli e rossi) e un gruppo eme (rosa) fino al coenzima Q
(beige chiaro), un trasportatore di elettroni mobile (dentro la membrana)
che porta gli elettroni al citocromo
bc1 (mdm 5-2011) nel complesso 3. L'enzima mostrato qui a destra
è batterico (file PDB 1nek).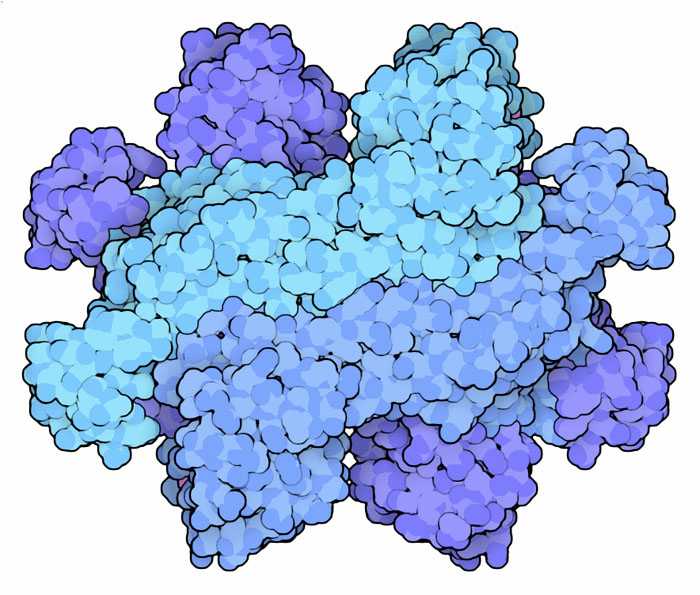
(7) Fumarasi La settima tappa è realizzata dall'enzima fumarasi che aggiunge acqua all'acido fumarico trasformandolo in acido malico. Qui a lato è mostrata una forma batterica di questo enzima (file PDB 1fuo). Nelle nostre cellule l'enzima si trova sia nei mitocondri, dove partecipa al ciclo dell'acido citrico, sia nel citoplasma, dove ha un ruolo attivo nella sintesi di molecole e anche (cosa sorprendente) nella risposta al danneggiamento del DNA. Le nostre cellule, però, hanno un solo gene che codifica per questo enzima, ma riescono a destinare alcuni enzimi ai mitocondri e altri al citoplasma utilizzando tempi diversi di ripiegamento della proteina. 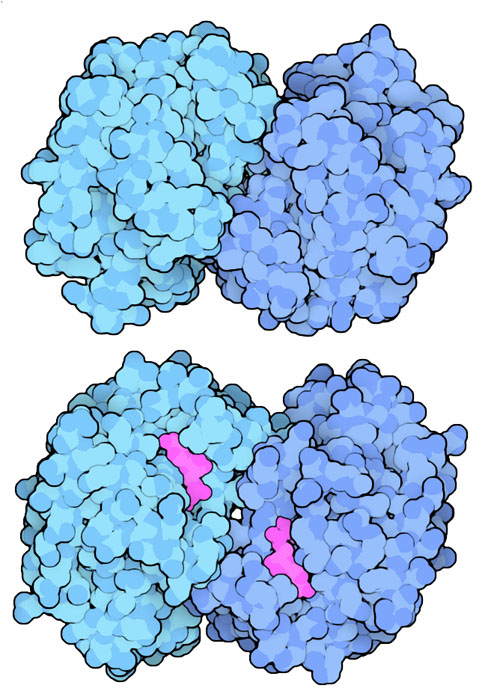 (8) Malato deidrogenasi L'ultima tappa del ciclo dell'acido citrico ricrea ossalacetato ossidando il gruppo alcolico dell'acido malico a chetone e trasferendo gli elettroni estratti al NADH. L'enzima malato deidrogenasi si trova sia nei mitocondri che nel citoplasma. Qui a destra sono mostrate le due forme, in alto quella mitocondriale (file PDB 1mld) e in basso quella citoplasmatica (file PDB 5mdh) nella quale sono ben visibili 2 molecole di NADH (magenta). Insieme queste due forme risolvono un difficile problema per la cellula. Il NADH prodotto nel citoplasma dalla glicolisi può essere smaltito in due modi. Può essere riconvertito in NAD+ con la fermentazione lattica, quella che negli sforzi intensi produce acido lattico nei nostri muscoli, oppure può essere mandato nei mitocondri per la reazione aerobica con l'ossigeno. La membrana dei mitocondri, però non può essere attraversata dal NADH, quindi entrano in gioco le due forme di malato deidrogenasi che agiscono da sistema navetta. Nel citoplasma il NADH è usato per ridurre ossalacetato a malato. Il malato può attraversare la membrana dei mitocondri e poi viene riconvertito in ossalacetato rigenerando NADH all'interno dei mitocondri. Spunti per ulteriori esplorazioni In molti casi gli enzimi del ciclo dell'acido citrico prodotti da organismi diversi sono molto simili tra loro. Usate lo strumento "Compare Structures" per valutare somiglianze e differenze tra le diverse forme che si trovano negli archivi del PDB. D. Voet and J. G. Voet (2011) Biochemistry, 4th Edition. John Wiley and Sons. O. Yogev, A. Naamati and O. Pines (2011) Fumarase: a paradigm of dual targeting and dual localized functions. FEBS Journal 278, 4230-4242. J. Rutter, D. R. Winge and J. D. Schiffman (2010) Succinate dehydrogenase -- assembly, regulation and role in human disease. Mitochondrion 10, 393-401. M. E. Fraser, K. Hayakawa, M. S. Hume, D. G. Ryan and E. R. Brownie (2006) Interactions of GTP with the ATP-grasp domain of GTP-specific succinyl-CoA synthetase. Journal of Biological Chemistry 281, 11058-11065. G. E. Murphy and G. J. Jensen (2005) Electron cytotomography of the E. coli pyruvate and 2-oxoglutarate dehydrogenase complexes. Structure 13, 1765-1773. P. Minarik, N. Tomaskova, M. Kollarova and M. Antalik (2002) Malate dehydrogenases -- structure and function. General Physiology and Biophysics 21, 257-265. |
||||||
|
|
||||||
