|
Molecola del Mese di Maggio 2014
Le aquaporine formano un canale che consente alle molecole d'acqua di
attraversare la membrana cellulare
 Introduzione Introduzione
Le membrane cellulari sono costituite da un doppio strato di fosfolipidi
e quindi sono una barriera quasi impenetrabile per le molecole
polari come l'acqua. Alcune cellule, però, devono consentire il
passaggio di acqua attraverso la membrana. Per esempio, la separazione
delle molecole di scarto nel rene e la pressione interna dell'occhio
richiedono un preciso controllo del flusso di acqua. In questi casi le
cellule usano dei particolari canali proteici di membrana, le acquaporine,
per controllare il flusso d'acqua dentro e fuori dalla cellula.
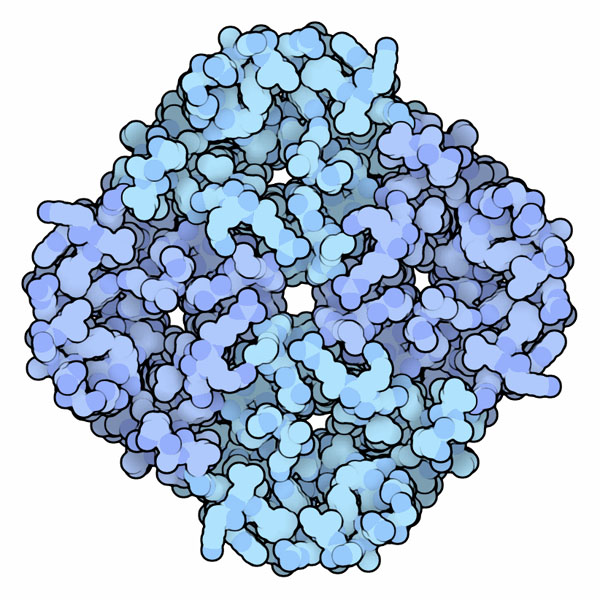
Quattro canali in uno
Le acquaporine si trovano in organismi molto diversi, dai batteri all'uomo,
ma hanno tutte una struttura molto simile. Questo significa che erano
presenti nei batteri che popolavano il pianeta Terra miliardi di anni
fa e che poi sono rimaste inalterate nel corso dell'evoluzione. Sono
formate da quattro catene proteiche identiche, ogni catena possiede
un piccolo canale al centro che consente il passaggio delle molecole
d'acqua una alla volta, in fila indiana. Il foro più grande che
si trova al centro delle quattro catene, invece, è circondato
da amminoacidi apolari e quindi non consente il passaggio di acqua.
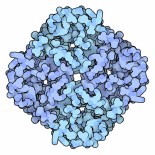
L'acquaporina mostrata qui a destra (file PDB 1fqy)
si tova nei globuli rossi del sangue.
Una recente struttura ad alta risoluzione (file PDB 3zoj),
non mostrata qui, ha rivelato che due amminoacidi carichi, all'ingresso
di ogni canale, sono importanti per assicurare che solo l'acqua neutra
possa attraversare il canale, ma non lo ione idrossonio H3O+
o lo ione idrossido OH-.
Controllare la acquaporine
Dato che le acquaporine sono essenziali per regolare l'osmosi in molti
tipi di cellule, gli scienziati stanno cercando un modo per bloccarne
l'azione nel tentativo di curare le malattie legate ai disturbi dell'equilibrio
idrico come il glaucoma. Le acquaporine sono state scoperte per
la prima volta osservando che il mercurio blocca il flusso di
acqua dentro e fuori dai globuli rossi, ma sfortunatamente il mercurio
è troppo velenoso per essere usato come farmaco. Ora gli scienziati
stanno cercando nuovi inibitori che possano essere usati come
farmaci per bloccare specifiche acquaporine.
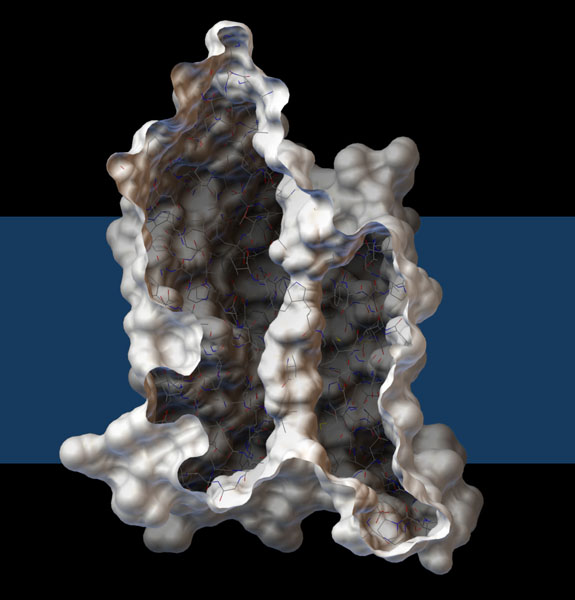
Acquagliceroporine
Nel nostro corpo sintetizziamo una dozzina di acquaporine, con funzioni
diverse, in vari tipi di cellule. Alcune acquaporine sono altamente
selettive e consentono il passaggio solo di acqua, mentre
altre possono lasciar passare anche altre molecole. Quella mostrata
qui a destra è una acquagliceroporina batterica (file PDB 1fx8)
che può far passare sia acqua che glicerina. Qui è
mostrata una sezione di una delle quattro subunità della
proteina che permette di vedere il canale (colonna bianca al
centro) che attraversa la membrana (fascia blu). Notate che c'è
una strozzatura nella parte superiore del canale, che controlla quali
molecole lo possono attraversare.
Esplorando la struttura
Il canale per l'acqua che attravera le acquaporine potrebbe causare
gravi problemi alle cellule. Una fila di molecole d'acqua, infatti,
può trasferire velocemente protoni da un lato all'altro
della membrana con una specie di passamano di protoni da una molecola
di acqua alla successiva. Questo sarebbe un disastro per la cellula
perchè potrebbe rapidamente dissipare il gradiente elettrochimico
che spinge la maggior parte delle pompe ioniche sulla superficie cellulare.
L'acquaporina riesce ad evitare questo problema impedendo alle molecole
d'acqua nel poro di avere tutte lo stesso orientamento e costringendole
ad orientarsi in due modi opposti sui due versanti del poro.
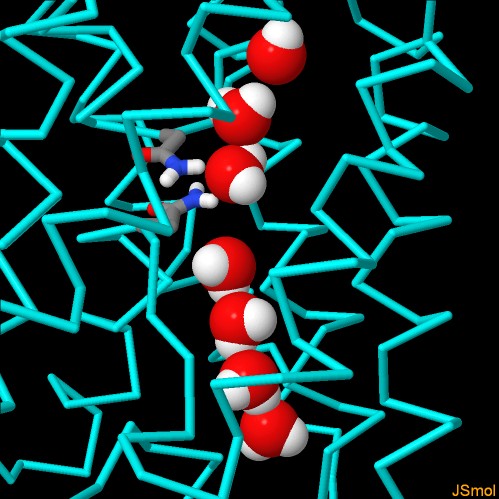
Questo è realizzato grazie a due amminoacidi di asparagina
(bastoncini colorati al centro dell'immagine qui sotto) che legano una
molecola d'acqua (sfera rossa e bianca) nella parte centrale del
canale e la costringono ad orientarsi in un certo modo. Questa molecola
d'acqua, insieme con gli altri amminoacidi lungo il canale, interagisce
con le altre molecole d'acqua del canale, allineate in fila indiana,
e le costringe a rivolgersi in direzioni opposte su due lati
del poro.
Nell'immagine qui sotto (file PDB 1ymg)
si possono vedere le molecole d'acqua in fila nel canale orientate in
modo opposto: le molecole d'acqua che enrano dall'alto rivolgono il
loro ossigeno (rosso) verso il centro del canale fino a quando interagiscono
con le due asparagine che le costringono a ruotare e procedono poi verso
il basso rivolgendo gli idrogeni bianchi verso l'uscita del canale.
(In un'altra struttura PDB 3zoj, non mostrata
qui, sembra che le molecole d'acqua legate alle due asparagine siano
due).
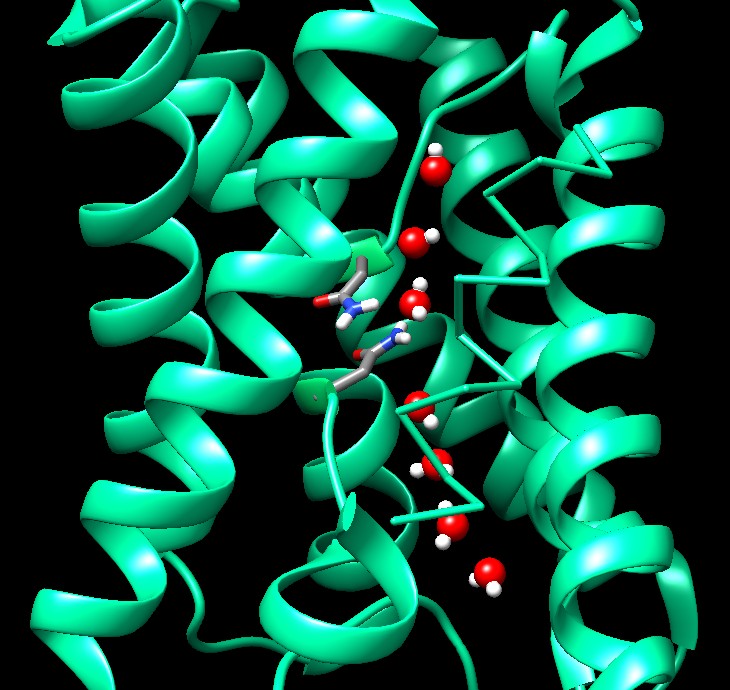
La stessa struttura (file PDB 1ymg) è
mostrata qui sotto elaborata con Chimera (vedi chimica
al computer).
. . . . . . . . . . . . . . . . 
Spunti per ulteriori esplorazioni
Negli archivi PDB ci sono le strutture di molte acquaporine diverse,
potete usare lo strumento "Compare Structures" (nel sito http://www.rcsb.org/pdb)
per esplorare le sequenze comuni.
Molte strutture di acquaporine hanno i canali chiusi da qualche amminoacido
che si è spostato per bloccare il canale dell'acqua. Gli scienziati
stanno ancora cercando di capire se questo fenomeno è dovuto
oppure no ad un meccanismo attivo di chiusura dei canali.
 Bibliografia Bibliografia
1fx8: D. Fu, A. Libson, L. J. Miercke, C. Weitzman, P. Nollert,
J. Krucinski & R. M. Stroud (2000) Structure of a glycerol-conducting
channel and the basis for its selectivity. Science 290,
481-486.
1fqy: K. Murata, K. Mitsuoka, T. Hirai, T. Walz, P. Agre, J.
B. Heymann, A. Engel & Y. Fujiyoshi (2000) Structural determinants
of water permiation through aquaporin-1. Nature 407, 599-605.
1ymg: W. E. C. Harries, D. Akhavan, L. J. W. Miercke, S. Khademi
& R. M. Stroud (2004) The channel architecture of aquaporin O at
2.2 Angstrom resolution. Proceedings of the National Academy of Science
USA 101, 14045-14050.
T. Gonen, T. Walz (2006) The structure of aquaporins. Quarterly
Reviews of Biophysics 39, 361-396.
A. S. Verkman (2012) Aquaporins in clinical medicine. Annual Review
of Medicine 63, 303-316.
3zoj: U. K. Eriksson, G. Fischer, R. Friemann, G. Enkavi, E.
Tajkhorshid & R. Neutze (2013) Subangstrom resolution X-ray structure
details aquaporin-water interactions. Science 340, 1346-1349.
A. S. Verkman, M. O. Anderson & M. C. Papadopoulos (2014)
Aquaporins: important but elusive drug targets. Nature Reviews Drug
Discovery 13, 259-277.
|
|
|
