|
|
Importine |
|||||
|
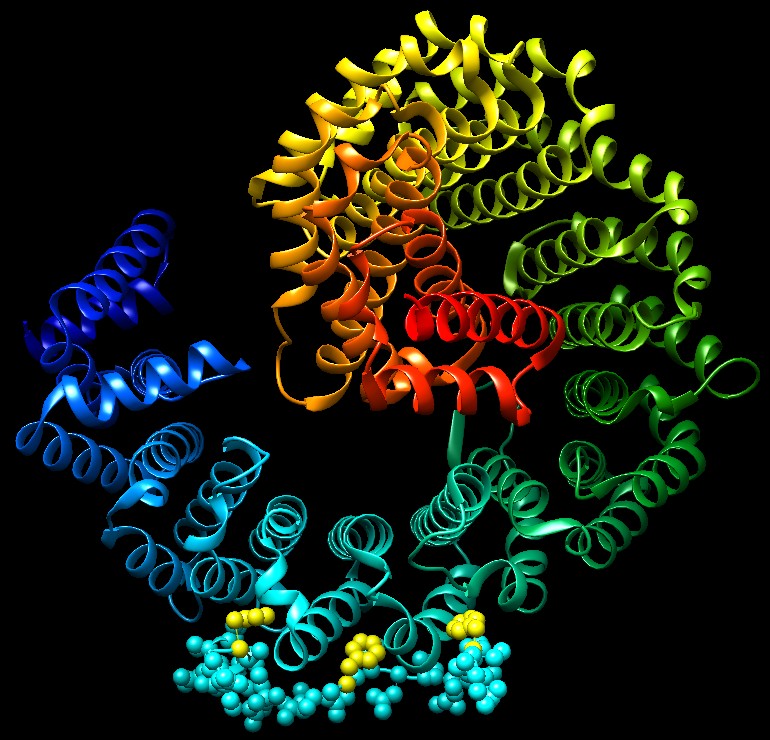
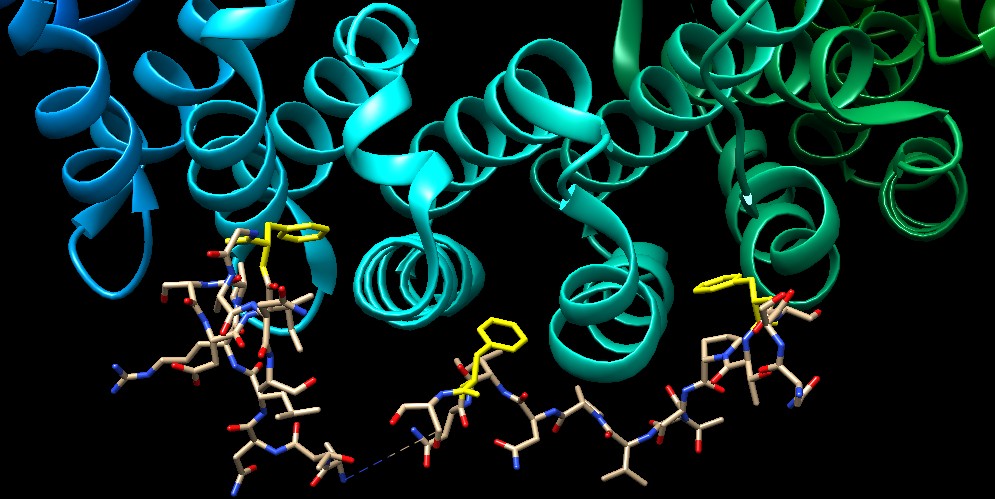
Molecola del Mese di Gennaio 2007 Le importine trasportano le proteine dal citoplasma al nucleo attraverso il complesso del poro nucleare 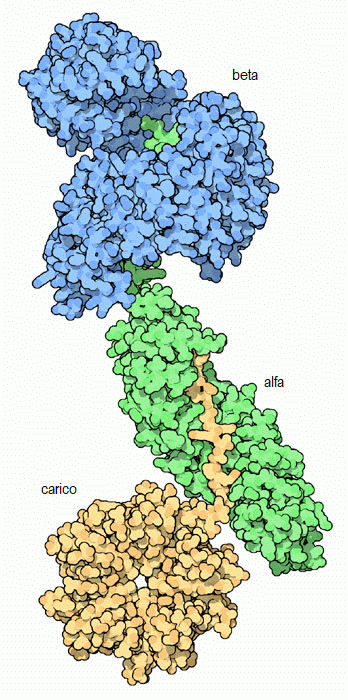 Introduzione IntroduzioneNelle nostre cellule il processo di sintesi delle proteine avviene in due zone ben distinte. La prima parte della sintesi, cioè la trascrizione del DNA per formare mRNA (mdm 1-2012), viene realizzata nel nucleo. La seconda parte, cioè la traduzione dell'mRNA in proteine, viene realizzata fuori dal nucleo, nel citoplasma, da organelli specializzati chiamati ribosomi (mdm 1-2010). Questa separazione richiede un continuo traffico di molecole: L'mRNA appena sintetizzato deve essere trasportato fuori dal nucleo e le proteine nucleari come istoni o RNA-polimerasi devono invece essere portate all'interno del nucleo. Nella membrana nucleare ci sono grandi pori a forma di tubo che agiscono come autostrade per collegare il nucleo con il citoplasma e ci sono poi delle proteine chiamate importine ed esportine (chiamate collettivamente carioferine) per trasportare le molecore avanti e indietro attraverso i pori. Trasporti speciali Le importine trasportano migliaia di proteine nel nucleo per realizzare compiti come compattare, leggere, copiare e riparare il genoma. Naturalmente, sarebbe troppo dispendioso costruire una importina diversa per ogni molecola da trasportare nel nucleo. Per questo molte proteine nucleari hanno una particolare etichetta, una corta sequenza di amminoacidi chiamata segnale di localizzazione nucleare, che informa il sistema di trasporto che quella proteina va portata nel nucleo. Le importine riconoscono questo segnale, si legano alla proteina e la trasportano attraverso il poro nucleare. Lavoro di importazione Il complesso dell'importina mostrato qui (ottenuto utilizzando tre diverse strutture PDB) trasporta proteine con segnali di localizzazione nucleare. Nella parte superiore, mostrata in blu, c'è l'importina beta (file PDB 1qgk). Si tratta del motore del complesso che riconosce i pori nucleari e si muove al loro interno. Si aggrappa all'estremità dell'importina alfa, mostrata al centro in verde (file PDB 1ee5). L'importina alfa è una molecola adattatrice che collega l'importina beta con il carico da trasportare. In questa figura, il carico è la nucleoplasmina, mostrata in basso in giallo (file PDB 1k5j) una proteina cheperon che aiuta l'avvolgimento del nucleosoma. Notate come il segnale di localizzazione nucleare del carico si estende verso l'alto ed è afferrato dall'importina alfa. Strategia d'uscita 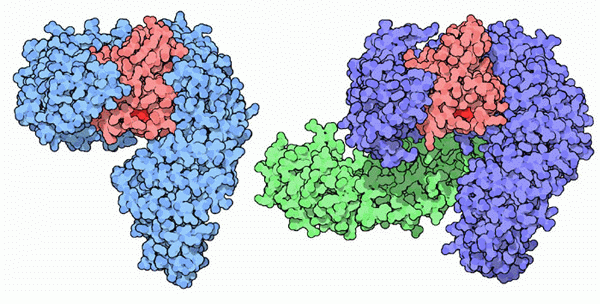 Dopo che il complesso importina beta - importina alfa - carico è entrato nel nucleo, il carico deve essere rilasciato e le importine devono essere rimandate indietro nel citoplasma per essere riutilizzate. La figura qui a fianco sulla sinistra mostra la proteina Ran (rossa) responsabile del rilascio del carico. Quando si lega all'importina beta (azzurra) ne cambia così tanto la forma da provocare il rilascio dell'importina alfa e del carico. Poi il complesso importina beta - Ran (qui a fianco sulla sinistra, file PDB 2bku) riattraversa il poro ed esce dal nucleo. Una volta nel citoplasma, la molecola di GTP legata all'interno di Ran (rosso vivo) viene scissa in GDP e fosfato. Questo provoca una variazione della forma di Ran che la fa staccare lasciando l'importina beta libera di portare nel nucleo un'altra proteina carico. L'importina alfa non è in grado di tornare da sola nel citoplasma, quindi si fa aiutare da un fattore nucleare di esportazione CAS (viola) mostrato qui sopra sulla destra (file PDB 1wa5). CAS è simile all'importina beta, ma si muove nel poro nucleare in direzione opposta. Si lega all'importina alfa e la trasporta fuori dal nucleo. Poi una scissione simile a quella del GTP in Ran rilascia l'importina alfa per un altro ciclo di trasporto. Esplorando la struttura Il reale meccanismo utilizzato dall'importina per trascinare le molecole attraverso i pori nucleari è ancora oggetto di dibattito, ma la struttura PDB 2bpt, rappresentata qui sotto, ci fornisce alcuni indizi su come potrebbe essere realizzato. Le migliaia di proteine che costituiscono il poro nucleare hanno in superfice speciali sequenze flessibili ricche di fenilalanina. Un lato dell'importina beta è in grado di aggrapparsi a queste sequenze speciali. La struttura mostrata qui include l'importina beta (colorata ad arcobaleno) e alcuni corti frammenti delle proteine del poro nucleare mostrate come piccole sfere (azzurre) in basso con le fenilalanine evidenziate in giallo. Notate che le fenilalanine si legano in tasche che si trovano tra le alfa eliche sulla superficie esterna dell'importina. L'importina beta può saltare da un posto al successivo nel poro nucleare aggrappandosi a queste particolari sequenze. . . . . . . . . . . . . . .
|
||||||
|
|
||||||