|
|
Recettori del glutammato |
|||||
|
Molecola del mese di luglio 2019 Nel cervello, i recettori per il glutammato, un neurotrasmettitore, sono di molte forme e dimensioni 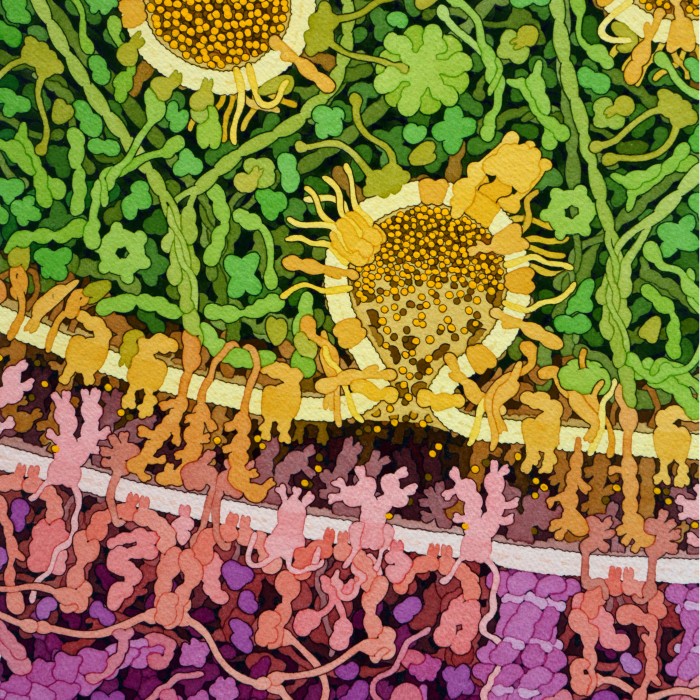 Introduzione IntroduzioneNel cervello, le cellule nervose comunicano tra loro usando piccole molecole di neurotrasmettitori. Questi sono di molte forme e dimensioni come l’acetilcolina (mdm 11/2005), la serotonina (mdm 8/2013) e le endorfine (piccoli peptidi di 5 amminoacidi, mdm 1/2018) e così consentono al sistema nervoso di variare il modo in cui i neuroni si parlano. Il glutammato, un semplice amminoacido, è il più comune neurotrasmettitore usato per trasportare messaggi eccitatori. Viene rilasciato dai neuroni nelle sinapsi dove ha il compito di eccitare il neurone successivo legandosi a specifici recettori per il glutammato, come si vede nell’illustrazione qui a lato di una sinapsi. In alto, pieno di corpuscoli verdi, vi è il neurone a monte che contiene vescicole colme di glutammato (sferette gialle). Quando il segnale nervoso giunge alla fine di questo neurone, viene trasmesso al neurone successivo (in basso, pieno di corpuscoli viola) grazie al rilascio del neurotrasmettitore glutammato che viene catturato da specifici recettori (rosa) infissi nella membrana del neurone a valle. Recettori AMPA del glutammato 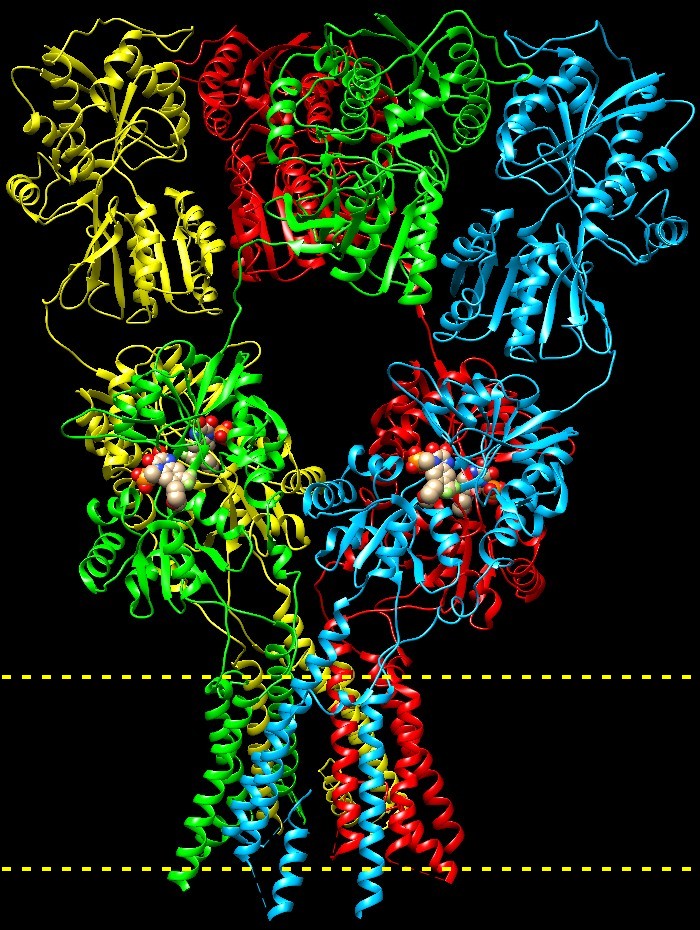 I
recettori AMPA (sensibili all’inibitore alfa-amino-3-idrossi-5-metil-4-isoxazolo
propionic acid) sono i più comuni recettori del glutammato nel
sistema nervoso centrale. Qui a fianco è mostrato un recettore
AMPA di topo (file PDB 3kg2). I
recettori AMPA (sensibili all’inibitore alfa-amino-3-idrossi-5-metil-4-isoxazolo
propionic acid) sono i più comuni recettori del glutammato nel
sistema nervoso centrale. Qui a fianco è mostrato un recettore
AMPA di topo (file PDB 3kg2). E' costituito da quattro catene proteiche con struttura modulare e ogni parte svolge un ruolo specifico. La porzione al centro riconosce e lega il glutammato che però, in questa struttura, è sostituito da un inibitore (sfere colorate). La porzione in basso è immersa nella membrana (indicata dai tratteggi) e forma un canale ionico che la attraversa costituito da 16 tratti ad alfa elica affiancati, 4 per ogni catena. Quando il glutammato si lega, genera un cambio di struttura che tira le catene del canale e lo fa aprire, permettendo agli ioni di fluire attraverso la membrana per giungere all’interno del neurone. Vi è anche una piccola coda sotto il canale ionico, visibile nella figura in alto. Qui a destra non è mostrata perche è flessibile e non è stata risolta ai raggi X. Interagisce con le proteine del citoscheletro che determinano la struttura della sinapsi. Apprendimento e memoria La comunicazione tra i neuroni è molto complessa, solo così può generare tutte le sfumature del pensiero e della memoria. I recettori AMPA hanno quattro catene, ognuna con un suo sito di legame per il glutammato e quindi il loro stato non è semplicemente acceso o spento. Inoltre i recettori AMPA, dopo essersi aperti in seguito alla ricezione del messaggio, si richiudono molto velocemente e diventano insensibili per un breve lasso di tempo. L'azione di immagazzinare dati di memoria può essere controllata in modo fine sia aggiungendo o togliendo recettori in una sinapsi, sia modificando alcuni amminoacidi in singoli recettori per modularne l’azione. 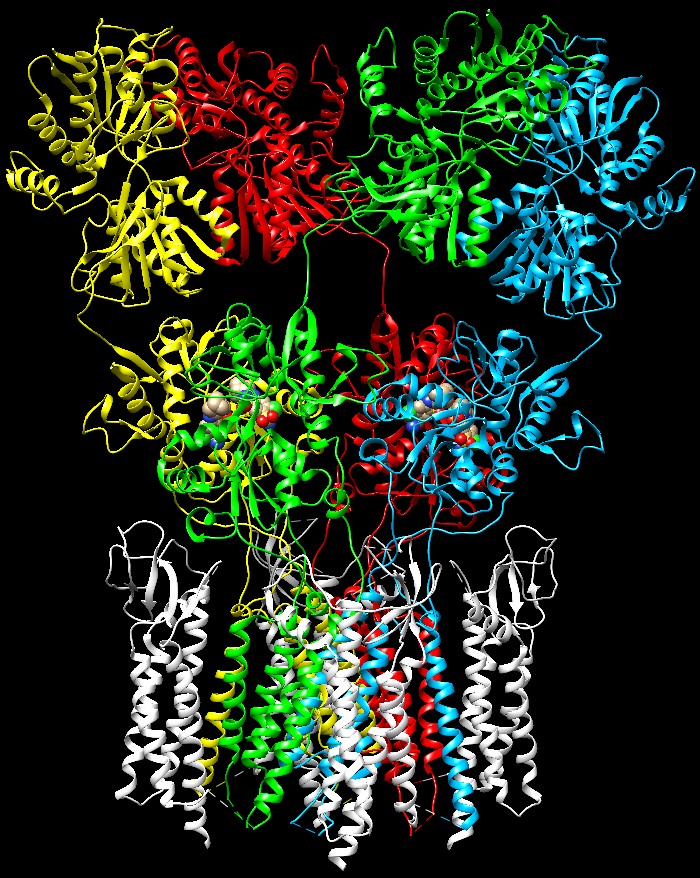 Livelli
di complessità Livelli
di complessitàPer rendere le cose ancora più complesse, il cervello ha ulteriori opzioni per modulare il segnale del glutammato. Alcune proteine accessorie si possono legare ai recettori AMPA come la proteina TARP mostrata qui a fianco (bianca) legata alla porzione che forma il canale ionico (file PDB 5weo). Inoltre ogni recettore è composto da combinazioni diverse di proteine simili. 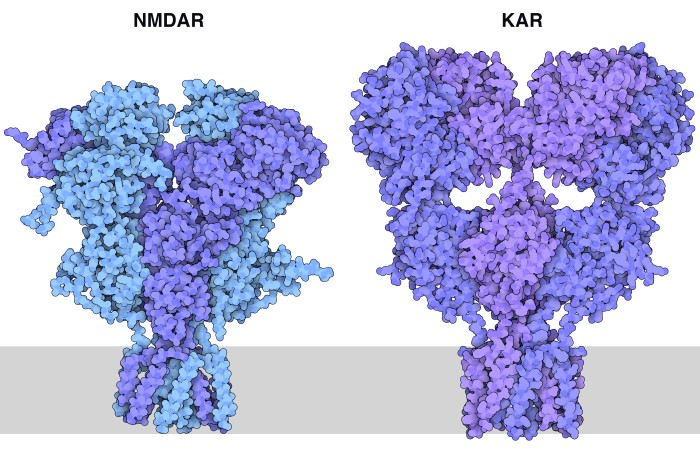 Infine vi sono altre due classi di recettori del glutammato che hanno azioni solo lievemente diverse. I recettori NMDA (file PDB 4pe5) e i recettori KA (kaianato, file PDB 5kuf) entrambi vengono chiamati così in base all’inibitore che li blocca e ne ha favorito la scoperta. L’insieme di questi recettori costituisce un sistema di segnalazione ricco di sfumature che ci consente di produrre i pensieri del nostro mondo interiore. 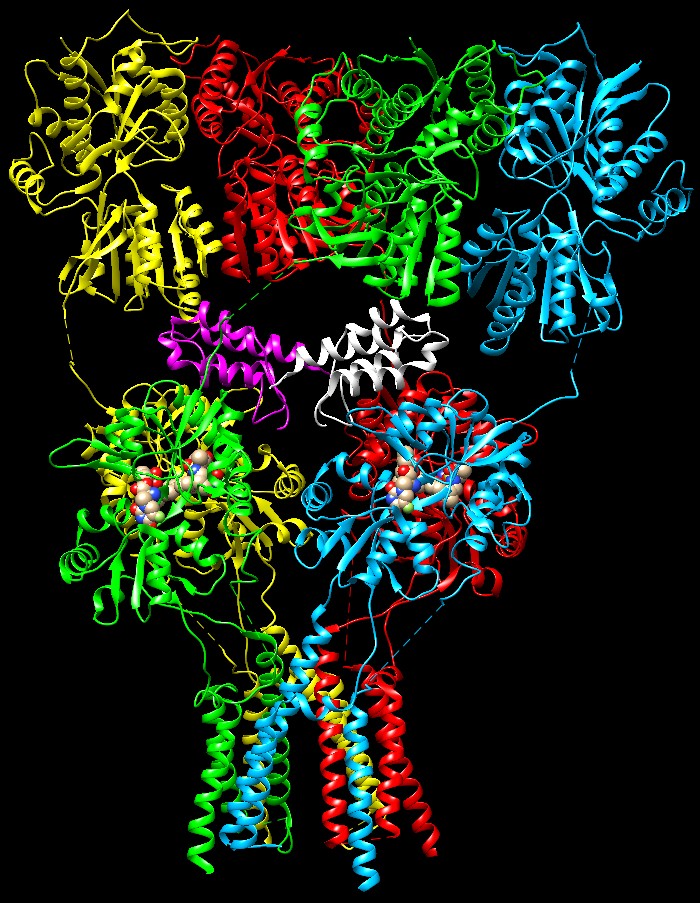 Esplorando
la struttura Esplorando
la strutturaAlcune conchiglie a cono producono una tossina estremamente velenosa costituita da una piccola proteina in grado di paralizzare le loro prede (in alcuni casi, è mortale anche per l'uomo). Questa ha come bersaglio i recettori del glutammato e dell'acetilcolina (mdm 11/2005). La struttura mostrata qui a fianco (file PDB 4u5c) mostra un recettore del glutammato avvelenato da questa tossina che lo ha bloccato nella posizione sempre aperta. Due copie di questa piccola proteina velenosa (una magenta e l'altra bianca) si sono legate al recettore inserendosi appena sopra la porzione che lega il glutammato e così la deformano in modo permanente. Questo induce le alfa eliche del canale ionico in basso ad aprirsi come il diaframma di una macchina fotografica. Spunti per ulteriori esplorazioni I recettori del glutammato sono molto flessibili. Se ne esaminate strutture diverse, noterete che hanno arrangiamenti diversi dei domini trans membrana rispetto ai domini che legano il glutammato. In tutte queste strutture mancano alcune porzioni del recettore che non sono state registrate a causa della grande flessibilità. Potete usare l’opzione Protein Feature View per determinare quale porzione della proteina è presente in ogni file. Chen, S., Gouaux, E. (2019) Structure and mechanism of AMPA receptor -- auxiliary protein complexes. Current Opinion in Structural Biology 54: 104-111. Zhu, S., Gouaux, E. (2017) Structure and symmetry inform gating principles of ionotropic glutamate receptors. Neuropharmacology 112: 11-15. 5weo: Twomey, E.C., Yelshanskaya, M.V., Grassucci, R.A., Frank, J., Sobolevsky, A.I. (2017) Channel opening and gating mechanism in AMPA-subtype glutamate receptors. Nature 549: 60-65. 5kuf: Meyerson, J.R., Chittori, S., Merk, A., Rao, P., Han, T.H., Serpe, M., Mayer, M.L., Subramaniam, S. (2016) Structural basis of kainate subtype glutamate receptor desensitization. Nature 537: 567-571. 4u5c: Chen, L., Durr, K.L., Gouaux, E. (2014) X-ray structures of AMPA receptor-cone snail toxin complexes illuminate activation mechanism. Science 345: 1021-1026. 4pe5: Karakas, E., Furukawa, H. (2014) Crystal structure of a heterotetrameric NMDA receptor ion channel. Science 344: 992-997. Yokoi, N., Fukata, M., Fukata, Y. (2012) Synaptic plasticity regulated by protein-protein interactions and posttranslational modifications. International Review of Cell and Molecular Biology 297:1-43. 3kg2: Sobolevsky, A.I., Rosconi, M.P., Gouaux, E. (2009) X-ray structure, symmetry and mechanism of an AMPA-subtype glutamate receptor. Nature 462: 745-756.
|
||||||
|
|
||||||
