|
|
Autofagia e |
|||||
|
Molecola del Mese di Novembre 2016 L'amminopeptidasi 1 con altri enzimi digestivi viene portata all'interno dei lisosomi della cellula per innescare il processo di autofagia 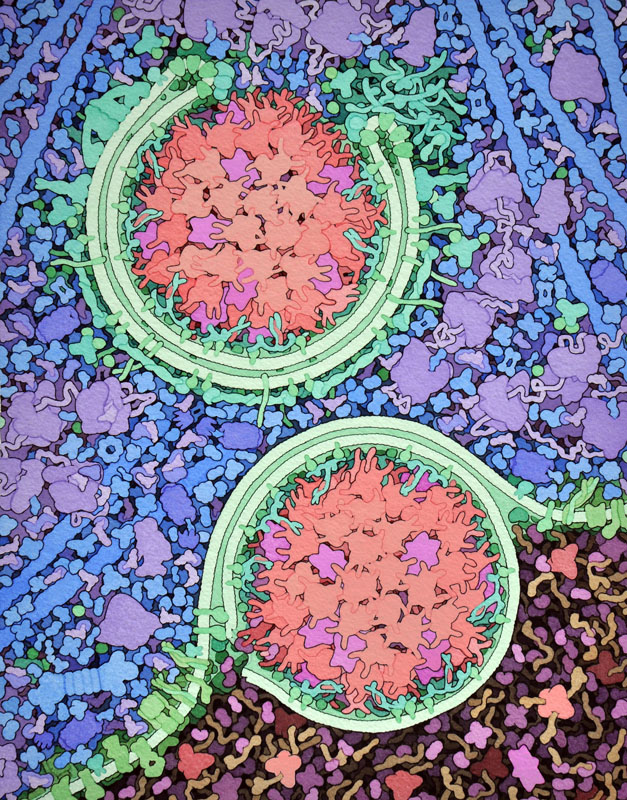 Introduzione IntroduzioneLe cellule devono continuamente sintetizzare nuove molecole, quando servono, e poi distruggerle quando hanno esaurito la loro funzione. Questa distruzione può avvenire nei proteasomi (mdm 8-2006) o negli esosomi (mdm 2-2007) organelli specializzati a degradare proteine ed RNA aggredendo una molecola per volta e riciclandone poi i componenti. Nelle cellule, però, vi è anche un altro meccanismo di distruzione, detto autofagia, che è in grado di rompere e poi riciclare le grandi biomolecole non una a una, ma in massa. Autofagia significa "mangiare se stessi" ed è proprio quello che fanno le cellule. Raccolgono i rifiuti e li portano nei lisosomi, organelli che costituiscono una specie di sistema digestivo interno alla cellula. Nella bella immagine qui a fianco ottenuta con la microscopia elettronica, si vede, in basso, uno spicchio di lisosoma in colori più scuri. Nel lisosoma le molecole vengono ridotte in pezzi da una miscela di enzimi digestivi che giungono sul posto trasportati in vescicole dette fagofori, mostrate qui a fianco circondate da una doppia membrana verde. Autofagia in azione L'autofagia ha molti ruoli importanti nelle cellule. Quando le cellule sono affamate, usano l'autofagia per mangiare e digerire parti del loro stesso citoplasma, e così possono sopravvivere nutrendosi con un auto cannibalismo. L'autofagia può anche essere più selettiva, circondando ed eliminando interi mitocondri o altri organelli danneggiati o diventati inutili. L'autofagia selettiva è anche usata in modo biosintetico, per portare enzimi digestivi dal citoplasma al lisosoma. I ricercatori si sono interessati all'autofagia per le sue connessioni con alcune malattie, per esempio, errori nell'autofagia sono collegati a malattie degenerative come l'Alzheimer mentre le cellule cancerose alterano l'autofagia per sopravvivere meglio. Dal citoplasma al vacuolo Il processo di autofagia selettiva è stato molto studiato nelle cellule di lievito, nelle quali un grande vacuolo al centro della cellula assume il ruolo di lisosoma. L'enzima amminopeptidasi 1 è portato nel vacuolo del lievito dai fagofori. Nella figura qui sopra in alto, si vede un fagoforo formato da una membrana a doppio strato (verde) che racchiude il suo carico di enzimi digestivi. In questo caso il carico è un insieme di enzimi come l'amminopeptidasi 1 (rosa) e l'alfa-mannosidasi (magenta). Più sotto, si vede il fagoforo in una fase successiva nella quale la sua membrana esterna si è fusa con la membrana del vacuolo, in attesa che delle lipasi nel vacuolo rompano la membrana interna rilasciando gli enzimi digestivi. Attivare l'amminopeptidasi 1 L'amminopeptidasi 1 è un enzima proteolitico che stacca uno alla volta gli amminoacidi di una proteina partendo dall'estremità ammino terminale. Altri enzimi proteolitici, come la carbossipeptidasi, fanno lo stesso partendo dall'estremità carbossi terminale. Naturalmente, vi sono anche altri enzimi digestivi che tagliano le proteine in mezzo alla loro catena come la pepsina (prodotta nello stomaco), la tripsina (mdm 10-203), la chimotripsina e l'elastasi (prodotte dal pancreas). 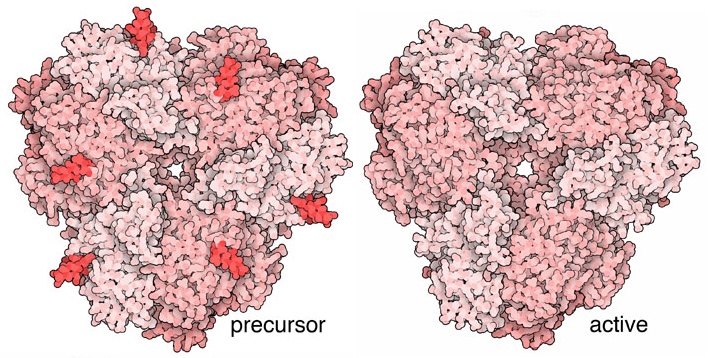 Come
altri enzimi proteolitici, anche l'amminopeptidasi 1 viene sintetizzata
in una forma inattiva, con un piccolo peptide di 45 amminoacidi
(in rosso) che sporge da un lato come si vede qui a fianco in questa grande
struttura formata da 12 catene proteiche (file PDB 5jh9).
Questo peptide interagisce con le molecole circostanti per formare aggregati
e interagisce anche con specifici recettori proteici dell'autofagia, in
questo modo si assicura che il fagoforo sia costruito subito attorno all'aggregato
inglobando solo una minima quantità di citoplasma. Come
altri enzimi proteolitici, anche l'amminopeptidasi 1 viene sintetizzata
in una forma inattiva, con un piccolo peptide di 45 amminoacidi
(in rosso) che sporge da un lato come si vede qui a fianco in questa grande
struttura formata da 12 catene proteiche (file PDB 5jh9).
Questo peptide interagisce con le molecole circostanti per formare aggregati
e interagisce anche con specifici recettori proteici dell'autofagia, in
questo modo si assicura che il fagoforo sia costruito subito attorno all'aggregato
inglobando solo una minima quantità di citoplasma. Solo quando l'enzima viene rilasciato dentro il vacuolo, il peptide, che costituisce la porzione ammino-terminale dell'enzima, viene eliminato. L'enzima si trasforma così nella forma attiva mostrata qui sulla destra (file PDB 4r8f) e può iniziare la sua azione distruttiva. 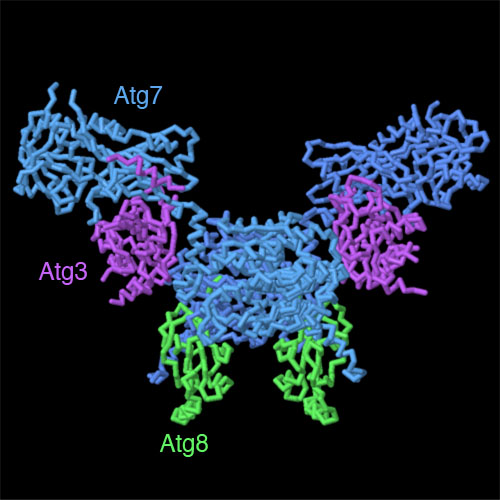 Esplorando la struttura
Esplorando la strutturaDecine di proteine contribuiscono alla costruzione del fagoforo e alla sua fusione con il lisosoma o con il vacuolo del lievito. Le proteine mostrate qui a destra attivano uno dei recettori proteici che trattengono l'amminopeptidasi 1 alla membrana del fagoforo (file PDB 4gsl in alto e 3rui in basso). Nel lievito, queste proteine vengono chiamate Atg, proteine connesse all'autofagia. In queste strutture Atg8 è la proteina trovata nella membrana del fagoforo, mentre Atg7 e Atg3 sono parte del macchinario che lo attiva attaccandogli una coda lipidica. Spunti per ulteriori esplorazioni Negli archivi PDB ci sono molte altre proteine correlate all'autofagia. Per esempio uno strano complesso a forma di S formato da molte proteine Atg (file PDB 4p1w) è coinvolto nel dare inizio al processo in caso di mancanza di cibo. Provate a fare una ricerca per “autophagy” nel sito PDB per vedere alcune di queste proteine. Nel 2016 Yoshinori Ohsumi ha vinto il premio Nobel per aver scoperto il meccanismo dell'autofagia. Cercate nel PDB il suo nome per trovare alcune delle strutture determinate nel suo laboratorio. L'enzima alfa-mannosidasi viene trasportato nel fagoforo insieme con amminopeptidasi 1. Esplorate la sua struttura nel file PDB 5jm0. Bibliografia H. Suzuki, T. Osawa, Y. Fujioka & N. N. Noda (2017) Structural biology of the core autophagy machinery. Current Opinion in Structural Biology 43, 10-17. X. Wen & D. J. Klionsky (2016) An overview of macroautophagy in yeast. Journal of Molecular Biology 428, 1681-1699. 5jh9: A. Yamasaki, Y. Watanabe, W. Adachi, K. Suzuki, K. Matoba, H. Kirisako, H. Kumeta, H. Nakatogawa, Y. Ohsumi, F. Inagaki & N. N. Noda (2016) Structural basis for receptor-mediated selective autophagy of aminopeptidase 1 aggregates. Cell Reports 16, 19-27. 4r8f: M. Y. Su, W. H. Peng, M. R. Ho, S. C. Su, Y. C. Chang, G. C. Chen & C. I. Chang (2015) Structure of yeast Ape1 and its role in autophagic vesicle formation. Autophagy 11, 1580-1593. 4gsl: S. E. Kaiser, K. Mao, A. M. Taherbhoy, S. Yu, J. L. Olszewski, D. M. Duda, I. Kurinov, A. Deng, T. D. Fenn, D. J. Klionsky & B. A. Schulman (2012) Noncanonical E2 recruitment by the autophagy E1 revealed by Atg7-Atg3 and Atg7-Atg10 structures. Nature Structural and Molecular Biology 19, 1242-1249. 3rui: S. B. Hong, B. W. Kim, K. E. Lee, S. W. Kim, H. Jeon, J. Kim & H. K. Song (2011) Insights into noncanonical E1 enzyme activation from the structure of autophagic E1 Atg7 with Atg8. Nature Structural and Molecular Biology 18, 1323-1330.
|
||||||
|
|
||||||
