|
Molecola del Mese di Luglio 2011
Le cellule aggiungono gruppi metilici al DNA per codificare ulteriori
informazioni dette epigenetiche che modificano l'espressione genica
 Introduzione Introduzione
Nel nostro corpo ci sono molti tipi di cellule
diverse, quelle della pelle, del sistema nervoso, delle ossa, ecc.
Queste hanno forma e dimensioni diverse, inoltre ogni tipo di cellula
costruisce un insieme diverso e caratteristico di proteine necessarie
per il suo funzionamento. Ogni cellula del nostro corpo, però,
ha lo stesso patrimonio genetico scritto nel suo DNA. Come fa ogni
cellula a sapere quali geni deve usare e quali invece deve ignorare?
Genetica ed epigenetica
Gli scienziati hanno scoperto che le informazioni
contenute nel DNA non si limitano alla semplice sequenza di basi azotate
che codificano il nostro patrimonio genetico. Le cellule sovrappongono
al loro patrimonio genetico ulteriori informazioni chiamate epigenetiche
che modificano l'espressione di alcuni geni. In qualche caso il controllo
si realizza per mezzo dei nucleosomi, cioè modificando gli istoni,
le proteine che si legano al DNA. In altri casi, le basi del DNA vengono
metilate, e così viene modificato il modo in cui vengono lette
durante la trascrizione impedendo quindi la sintesi proteica.
Tabula rasa
Nei primi minuti di vita, quando siamo costituiti
da una sola cellula, l'informazione epigenetica è azzerata.
Nell'uovo fecondato i gruppi metilici sono stati rimossi e tutti i geni
sono sullo stesso piano. Poi, quando le cellule si dividono formando
l'embrione, devono differenziarsi e diventare cellule della pelle, del
sistema nervoso, o quant'altro. A questo punto entrano in funzione enzimi
chiamati DNA metilasi che aggiungono gruppi metile ai geni e così
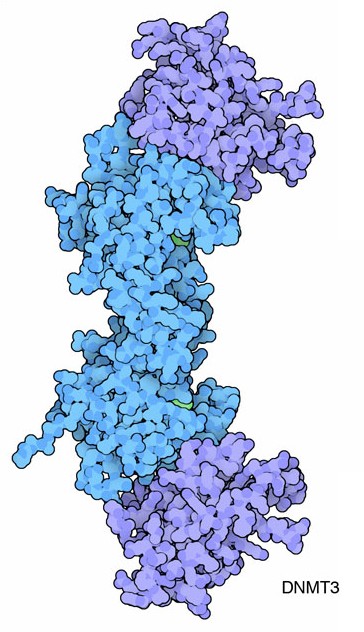
ne spengono alcuni e ne attivano altri. La DNA metilasi DNMT3, mostrata
qui dal file PDB 2qrv, realizza il suo
importante lavoro creando il giusto codice epigenetico di gruppi metilici
lungo il genoma.
 Manutenzione dei metili
Manutenzione dei metili
Dopo che ogni cellula si è differenziata,
il codice epigenetico deve essere mantenuto per il resto della vita
dell'organismo. Quando una cellula si divide, l'informazione deve essere
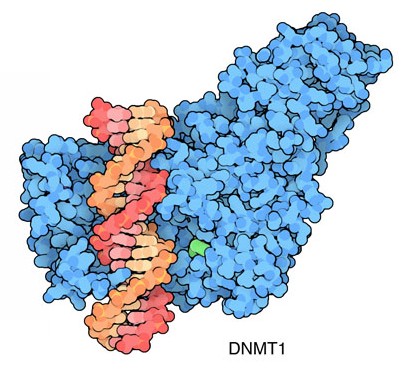
trasmessa alle nuove cellule. La DNA metilasi DNMT1, mostrata qui a
destra dal file PDB 3pt6, realizza questo
lavoro. Quando il DNA viene duplicato, aggiunge i gruppi metilici alle
nuove catene. L'informazione viene trasmessa grazie ad un espediente:
i gruppi metilici vengono quasi sempre aggiunti alle basi di citosina
che appartengono alla seguente sequenza palindroma (che si
legge nello stesso modo, in direzioni opposte, nei due filamenti):
. . . . . . . . . . . . . . . . . . . . . . .
-------CpG-------
. . . . . . . . . . . . . . . . . . . . . . .
-------GpC-------
In questa coppia di basi la citosina è presente su tutte e due
le catene così, in un tratto di DNA metilato, tutte e due le
catene hanno un gruppo metilico. Quando il DNA viene duplicato, le due
doppie eliche figlie sono composte da una catena vecchia, metilata
in modo corretto, e da una nuova, non metilata. L'enzima DNMT1
ha il compito di cercare le sequenze CpG che sono metilate su una
sola catena e quindi metila anche l'altra catena.
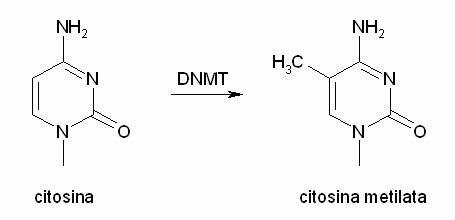
Citosina metilata
La metilazione della citosina avviene nella
posizione 5', questo altera il profilo della molecola nella parte sinistra,
però non cambia la parte destra della molecola che è implicata
nell'appaiamento con la guanina con la quale forma tre legami idrogeno
che tengono uniti i due filamenti nella doppia elica del DNA.
. . . . . . . . . . . . . . . . . . . . . . .
. . . . 
Nel DNA, il gruppo metile sporge nella fenditura principale e quindi
la citosima metilata può essere riconosciuta toccando lateralmente
la catena del DNA senza bisogno di separare le due catene.
 Batteri
restrittivi Batteri
restrittivi
Anche i batteri metilano il DNA, però non
lo fanno per accendere o spegnere geni specifici, ma per difendersi
dai virus. I batteri, infatti, sintetizzano gli enzimi
di restrizione (mdm 8-2000) che sono specializzati nel tagliare
il DNA dei virus in corrispondenza di specifiche sequenze di basi azotate.
I batteri, naturalmente, devono proteggere il proprio DNA dal rischio
di essere tagliato dai propri enzimi di restrizione e quindi sintetizzano
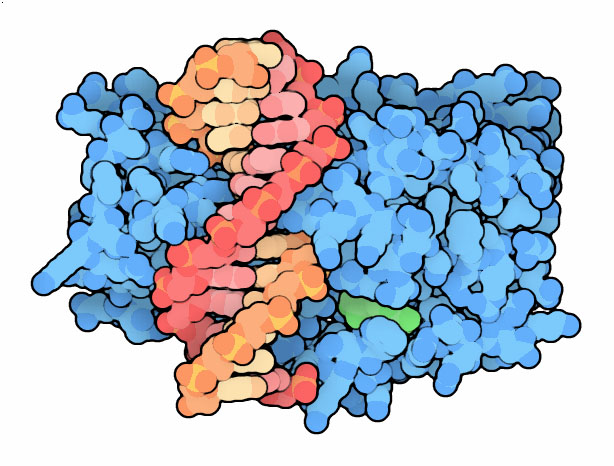
delle specifiche DNA metilasi, come quella mostrata qui a destra, Hha1(file
PDB 1mht), che aggiungono gruppi metilici
al loro DNA in corrispondenza di quelle stesse sequenze. I gruppi metilici
impediscono agli enzimi di restrizione di riconoscere e tagliare quella
sequenza, ma consentono comunque al DNA di funzionare correttamente,
cioè le basi metilate possono essere lette durante le operazioni
di duplicazione e trascrizione. In questo modo gli enzimi di restrizione
si muovono nella cellula batterica senza fare nulla fino a quando un
virus infetta la cellula. Il DNA virale non è metilato e quindi
viene rapidamente tagliato in pezzi dagli enzimi di restrizione. Questo
meccanismo può essere interpretato come una specie di sistema
immunitario primitivo.
Esplorando la struttura
E' stato necessario studiare la struttura di molte
DNA metilasi per capire i vari aspetti del suo funzionamento.
Qui sotto sulla sinistra è mostrato l'enzima DMNT1 legato
ad un piccolo pezzo di DNA (file PDB 3pt6).
L'enzima si avvolge intorno al DNA ed esplora i bordi delle basi azotate
nella fenditura principale per vedere se sono metilate, usando la porzione
di alfa elica mostrata in azzurro. Questa struttura contiene un DNA
privo di gruppi metilici, che normalmente non viene modificato dalla
DNA metilasi. L'enzima riconosce che le basi non sono metilate nè
su una catena nè sull'altra e tiene quindi il DNA lontano dal
sito attivo.
. . . . . . . . . .
. . . . . .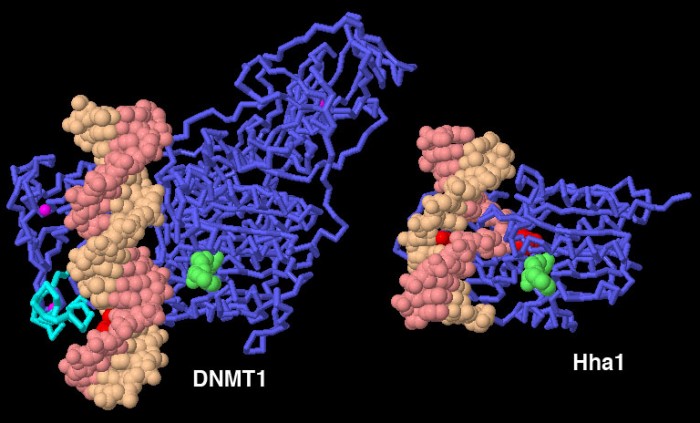
Finora non è stata risolta nessuna struttura di DNMT1 che lega
DNA metilato su una sola catena, ma possiamo farci un'idea di come si
comporterebbe l'enzima in questo caso, osservando una metilasi batterica,
Hha1 (file PDB 1mht) mostrata qui
sopra a destra e, più in dettaglio, nell'immagine qui sotto.
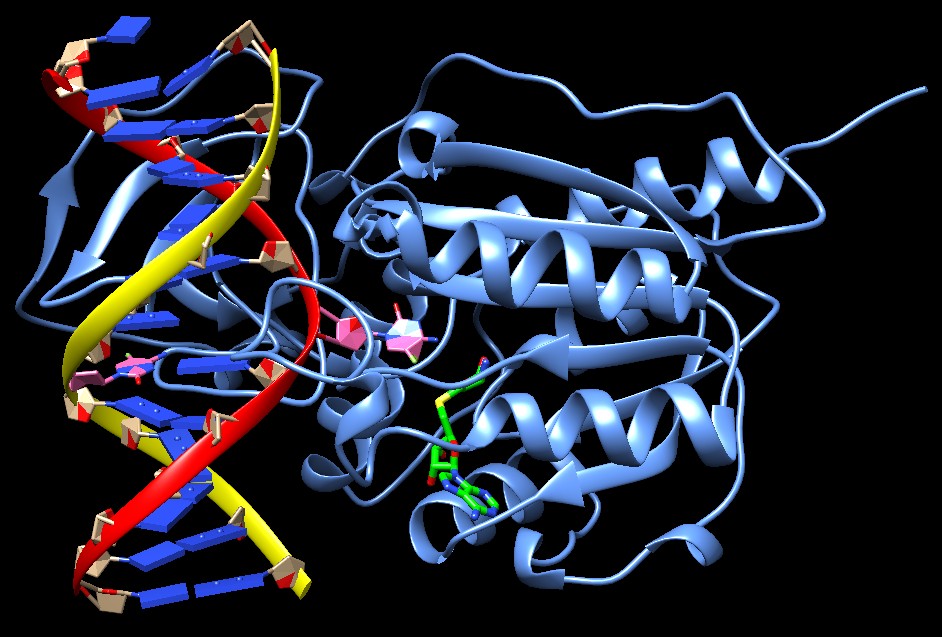
Il DNA è legato in una tasca profonda dell'enzima ed ha la normale
struttura a doppia elica ad eccezione della citosina (rossa sopra,
rosa qui sotto) che deve essere metilata. Questa infatti è stata
piegata verso l'esterno della catena rossa e così
si viene a trovare nel sito attivo a contatto con la molecola (verde)
di S-adenosilmetionina che le deve donare il gruppo metile (in questa
struttura sostituita con un analogo non reattivo).
Notate, qui sotto, che nella catena gialla del DNA c'è l'altra
citosina (della sequenza palindroma) già metilata, anche questa
è mostrata in rosa.
Questa immagine è stata creata con Chimera (vedi chimica
al computer).
. . . .
Spunti per ulteriori esplorazioni
1. Il gruppo metilico trasferito alla citosina
dalla DNA metilasi viene fornito dal coenzima S-adenosilmetionina.
Molte strutture di DNA metilasi sono state determinate usando al posto
del normale coenzima una molecola simile, ma non reattiva, S-adenosyl
homocysteine, che si comporta quindi da inibitore competitivo. Alcune
strutture, come PDB 3av6, usano un inibitore
diverso, chiamato SAM. Cercate negli archivi PDB altri enzimi che contengono
questo legando. Fanno reazioni simili?
2. Confrontate la struttura di enzimi DNMT con altre metilasi
batteriche usando lo strumento "structure comparison" nel
sito PDB. Notate che il nucleo di tutti questi enzimi è molto
simile, ma gli enzimi DNMT sono molto più grandi. Quale credete
sia il motivo per cui le DNA metilasi hanno bisogno di ulteriori domini
oltre a quello che realizza la reazione?
 Bibliografia Bibliografia
R. Z. Jurkowska,
T. P. Jurkowski and A. Jeltsch (2011) Structure and Function of Mammalian
DNA Methyltransferases. ChemBioChem 12, 206-222.
X. Cheng, R. M. Blumenthal (2008) Mammalian DNA Methyltransferases:
A Structural Perspective. Structure 16, 341-350.
|
|
|
