|
|
Canali meccanosensibili |
|||||
|
Molecola del Mese di Gennaio 2008 I canali sensibili alla pressione si aprono quando la pressione interna alla cellula diventa troppo alta 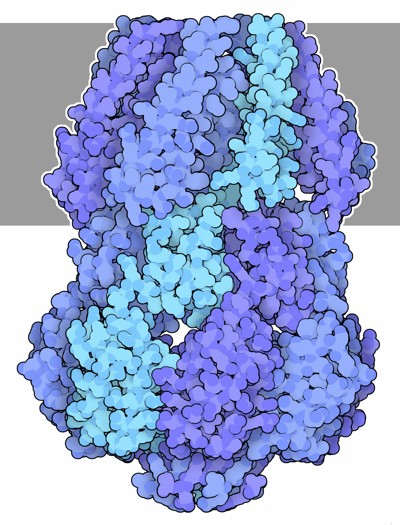 Introduzione IntroduzioneNoi siamo molto resistenti ai cambiamenti dell'ambiente che ci circonda. Il nostro corpo ha dimensioni così grandi che possiamo resistere a condizioni climatiche estreme di caldo e freddo, inoltre la nostra pelle ci protegge quando facciamo un bagno in acqua dolce o salata. Se poi le condizioni ambientali diventano troppo sfavorevoli, possiamo allontanarci e cercare un posto meno caldo, meno freddo o più asciutto. I batteri non possono fare altrettanto. Sono molto piccoli e vivono immersi in acqua, quindi i cambiamenti ambientali per loro costituiscono un grave problema. Per esempio, se piove, si possono trovare all'improvviso immersi in acqua piovana, molto povera di sali, in queste condizioni l'acqua può entrare per osmosi nelle loro cellule e ne fa aumentare la pressione interna fino a farli esplodere. Può anche accadere che i batteri si trovino improvvisamente immersi in acqua salata, questa fa uscire acqua dalle loro cellule per osmosi e li disidrata. I batteri hanno sviluppato delle strategie per resistere a questi stress ambientali e così riescono a mantenere al loro interno una giusta pressione osmotica. Sotto pressione I batteri controllano la loro pressione interna cambiando la concentrazione delle molecole al loro interno. Se si trovano in acqua salata, resistono alla disidratazione trasportando ioni al loro interno e sintetizzando altre piccole molecole. D'altra parte, se vengono a trovarsi in acqua povera di sali, riducono la loro concentrazione interna di sali e piccole molecole. In questo caso, usano dei canali meccanosensibili che si aprono e permettono alle loro piccole molecole di uscire. Le cellule batteriche poste in acqua distillata, perdono più del 95% delle loro piccole molecole come amminoacidi, zuccheri e ioni potassio. Conservano però al sicuro, al loro interno, proteine e ribosomi e quindi possono recuperare velocemente la piena efficienza iniziando la sintesi proteica pochi minuti dopo che le condizioni ambientali sono tornate normali. Valvole a pressione I canali meccanosensibili, rispondono alla pressione interna della cellula aprendosi quando questa raggiunge livelli pericolosi. Avvertono la pressione sentendo i cambiamenti delle proprietà della membrana. Quando la membrana è stirata dalla forte pressione interna, provoca un cambiamento conformazionale che apre il canale. La cellula possiede molti canali che si aprono a differenti livelli di pressione. Il canale MscS, mostrato qui sopra (file PDB 2oau), si apre a livelli di pressione moderati, mentre quello mostrato più sotto si apre a pressioni più elevate. Canale piccolo Il canale meccanosensibile a piccola conducibilità MscS è composto di sette catene identiche legate tra loro per formare le pareti di un tubo. Il canale attraversa la membrana cellulare e forma un poro sensibile alla pressione, con un diametro di circa 13 Angstrom. Questo è così piccolo che consente il passaggio solo di piccole molecole e di ioni. La porzione che sporge nel citoplasma (nella figura qui sopra si trova in basso) forma una cupola di forma sferica con un foro al centro della volta creando così una specie di anticamera che limita le dimensioni delle molecole che possono raggiungere il poro quando si apre. Canal grande Il canale meccanosensibile a grande conducibilità MscL mostrato qui sotto (file PDB 2oar) entra in azione nelle vere emergenze, quando la pressione diventa così elevata che la cellula può scoppiare. Quando si apre, forma un poro di dimensioni maggiori rispetto al canale MscS, con un diametro di 30-40 Angstrom. E' formato da cinque catene identiche che si aprono come il diaframma di una macchina fotografica quando la membrana cellulare è fortemente stirata. La struttura mostrata qui sotto è nella forma chiusa. . . . . . . . . . . .  Esplorando la struttura Potete vedere il canale MscS in azione in due file PDB: la forma chiusa è illustrata qui sotto sulla sinistra (file PDB 2oau), la forma aperta è mostrata qui sotto sulla destra (file PDB 2vv5). In entrambe le strutture la visuale è lungo l'asse del poro. Due amminoacidi di leucina (bianchi) in ognuna delle sette catene formano una barriera circolare che è in grado di bloccare il passaggio attraverso il canale. Potrebbe sembrare che l'anello di amminoacidi non sia abbastanza chiuso e che possa permettere il passaggio di piccole molecole, ma dato che le leucine sono amminoacidi apolari, quindi idrofobici, impediscono il passaggio delle molecole di acqua. Questo anello viene chiamato a "sigillo di vapore" per la sua resistenza a farsi bagnare dall'acqua. 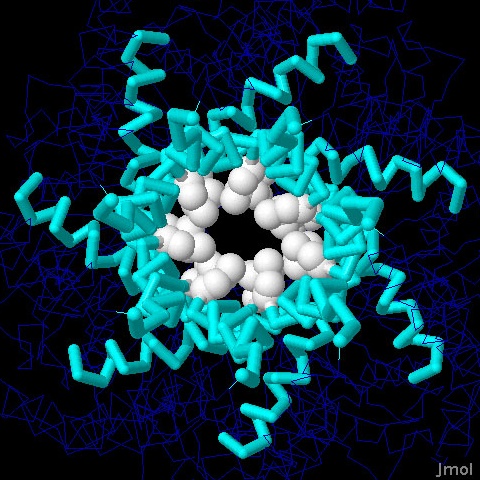 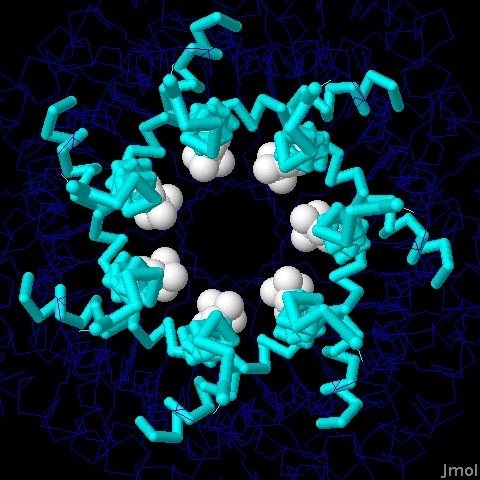 Nella figura qui sotto a destra è illustrato il canale MscS nella forma chiusa (file PDB 2oau) visto di lato. Nella parte alta si vede la porzione flessibile della proteina che costituisce il poro. E composta da tratti avvolti ad alfa elica (rossi), questi sono mobili e consentono al poro di allargarsi quando la tensione della membrana sale e tira queste catene verso l'esterno. La parte inferiore è la porzione rigida che costituisce l'anticamera a forma di cupola sferica che sporge all'interno della cellula nella quale si possono accumulare solo le piccole molecole e gli ioni che riescono a passare dal piccolo foro di diametro fisso posto in basso. Nella figura qui sotto a sinistra è mostrato il poro visto dal basso. Da questa immagine possiamo capire meglio la struttura del poro formato da sette catene proteiche assemblate in modo stretto per formare una perfetta macchina molecolare. La parte inferiore del poro è composta prevalentemente da tratti beta pieghe (gialli) che sono più rigidi grazie ai legami idrogeno che legano una all'altra le catene e le tengono affiancate. 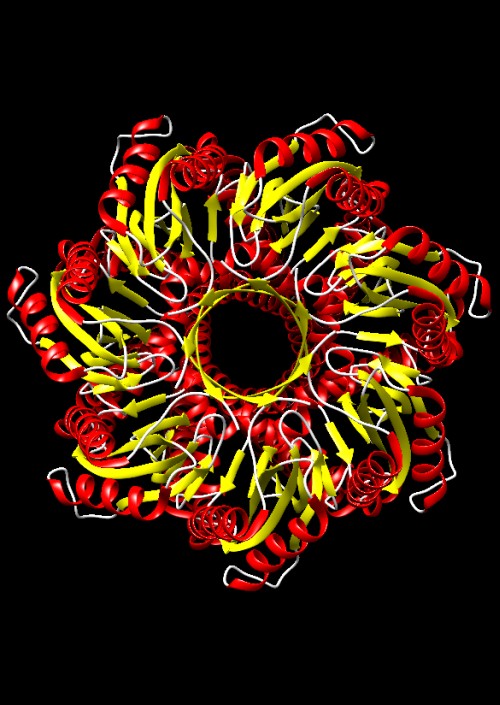 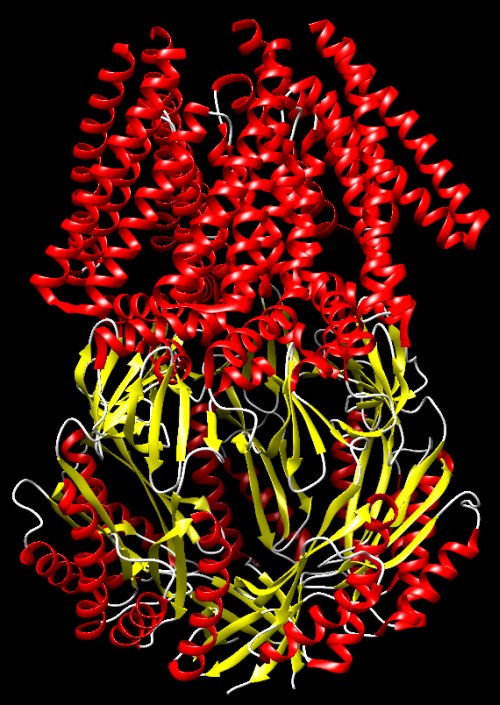 Spunti per ulteriori esplorazioni 1-- Esistono molti altri canali di membrana composti di più catene proteiche identiche assemblate in una struttura a tubo. Provate a cercarne qualche esempio negli archivi PDB. 2-- I canali per il potassio sono altamente specifici, permettono solo il passaggio di ioni potassio, mentre i canali meccanosensibili permettono il passaggio di molti tipi di ioni e di piccole molecole. Quali particolari, in queste strutture, producono caratteristiche funzionali così diverse? C.S. Gandhi, D.C.Rees (2008) Opening the molecular floodgates. Science 321, 1166-1167. I.R. Booth, M.D. Edwards, S. Black, U. Schumann and S. Miller (2007) Mechanosensitive channels in bacteria: signs of closure? Nature Reviews Microbiology 5, 431-440. A. Anishkin, C. Kung (2005) Microbial mechanosensation. Current Opinion in Neurobiology 15, 397-405. C. Kung (2005) A possible unifying principle for mechanosensation. Nature 436, 647-654. E. Perozo, D. C. Rees (2003) Structure and mechanism in prokaryotic mechanosensitive channels. Current Opinion in Structural Biology 13, 432-442.
|
||||||
|
|
||||||
