|
Molecola del mese di maggio 2003
L'emoglobina cambia leggermente forma quando
si carica e si scarica di ossigeno in modo da aumentare la sua efficienza
 Sangue rosso, sangue blu Sangue rosso, sangue blu
Il sangue ossigenato è rosso brillante mentre quello deossigenato,
ha un colore rosso scuro, ma appare blu quando scorre attraverso le vene,
specialmente nelle persone con pelle chiara. Questo è dovuto al
modo in cui i diversi colori della luce viaggiano attraverso la pelle:
la luce blu è riflessa dagli strati superficiali della pelle, mentre
la luce rossa penetra più profondamente. Alcuni organismi come
le lumache e i granchi, d'altra parte, usano il rame e non il ferro per
trasportare l'ossigeno, così hanno veramente il sangue blu.
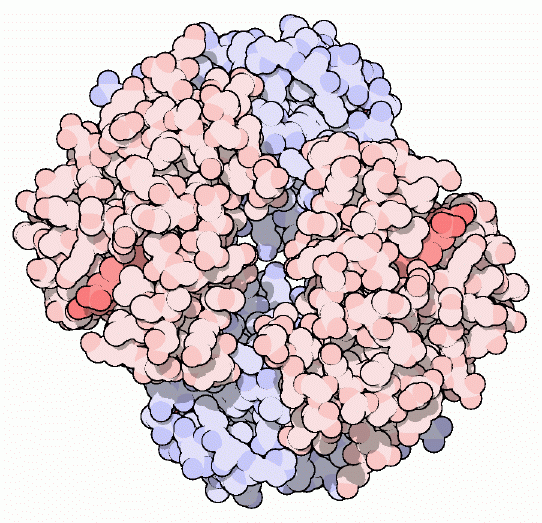
L'emoglobina è la proteina che rende rosso il sangue. È
composta di quattro catene proteiche, due catene alfa e due catene
beta, ognuna con un eme a forma di anello che contiene un atomo
di ferro al centro. L'ossigeno si lega in modo reversibile agli atomi
di ferro e così viene trasportato attraverso il sangue.
Ognuna delle quattro catene dell'emoglobina ha una
struttura simile alla mioglobina
(mdm 1-2000), la proteina usata per immagazzinare l'ossigeno nei muscoli
e in altri tessuti. La struttura complessa dell'emoglobina formata da
quattro catene è essenziale per il suo funzionamento, perchè
le consente di cambiare forma e affinità per l'ossigeno a seconda
della quantità di ossigeno presente.
Uso e abuso dell'emoglobina
Oltre a trasportare ossigeno, l'emoglobina può
legare e trasportare anche altre molecole come l'ossido nitrico e il
monossido di carbonio.
L'ossido nitrico NO è attivo sulle pareti dei vasi
sanguigni e ne causa il rilassamento. Questo fatto ha come conseguenza
una riduzione della pressione del sangue. Studi recenti hanno dimostrato
che l'ossido nitrico si può legare a specifici residui di cisteina
formando emoglobina
S-nitrosilata (mdm 5/2019) ed anche agli atomi di ferro nei
gruppi eme. Così, l'emoglobina contribuisce alla regolazione
della pressione sanguigna distribuendo ossido nitrico attraverso il
sangue.
Il monossido di carbonio CO, invece, è un gas tossico
che sostituisce rapidamente l'ossigeno nei gruppi eme, formando complessi
stabili dai quali è difficile da rimuovere. Questo abuso dei
gruppi eme impedisce all'ossigeno di legarsi al ferro per essere trasportato
ai tessuti, e può portare alla morte per soffocamento.
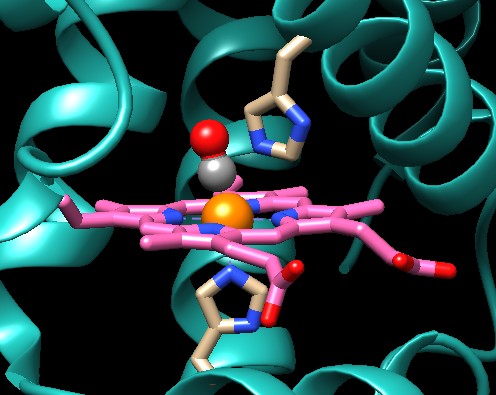
Come è mostrato qui sotto a sinistra (file PDB 2hco)
il monossido di carbonio (sfere grigia e rossa) si lega in modo leggermente
obliquo sul ferro dell'eme, ma farebbe un legame molto più forte
se potesse legarsi perpendicolarmente. E' costretto a legarsi obliquamente
a causa dell'ingombro di un amminoacido di istidina posto subito sopra
il ferro dell'eme. Questa istidina rappresenta una difesa per l'emoglobina
che altrimenti sarebbe avvelenata anche da minime concentrazioni di
monossido di carbonio.
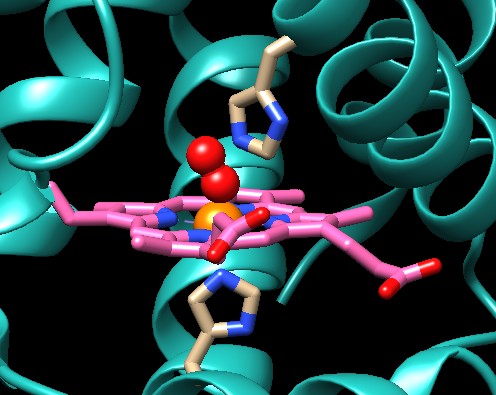
Nell'immagine qui sopra sulla destra (file PDB 1hho),
invece, si vede che l'ossigeno O2
(due sfere rosse) si lega obliquamente sul ferro, ma questo è
il suo legame spontaneo e non viene ostacolato dall'istidina.
Sangue artificiale
Le trasfusioni di sangue hanno salvato innumerevoli
vite. Comunque, il bisogno di sangue del giusto tipo, la breve durata
del sangue immagazzinato, e la possibilità di contaminazione
sono ancora dei gravi problemi. La comprensione di come funziona l'emoglobina,
basata su decenni di studi e su molte strutture cristallografiche, ha
stimolato la ricerca sul sangue artificiale e su sostituti del sangue.
L'approccio più ovvio è usare una soluzione di emoglobina
pura per sostituire il sangue perduto. Il problema principale è
tenere insieme le quattro catene proteiche dell'emoglobina. Senza
il guscio protettivo dei globuli rossi le quattro catene si separano
rapidamente. Per evitare questo problema sono state progettate molecole
di emoglobina insolite, dove due delle quattro catene sono legate tra
loro chimicamente, come mostrato nel file PDB 1c7d.
In questa struttura, due residui supplementari di glicina formano un
collegamento tra due catene, impedendo la loro separazione in soluzione.
Cugini dell'emoglobina
Negli archivi del PDB, ci sono molte molecole di
emoglobina. Potete trovare la struttura pionieristica di Max Perutz
dell'emoglobina di cavallo mostrata nella figura all'inizio della pagina
(file PDB 2dhb). Ci sono strutture di
emoglobina umana, sia di adulto che fetali. Potete trovare anche emoglobine
insolite come la leghemoglobina che è stata trovata nei
legumi. Si pensa che protegga i batteri ossigeno-sensibili che fissano
l'azoto nelle radici delle piante di leguminosa. Da non molto è
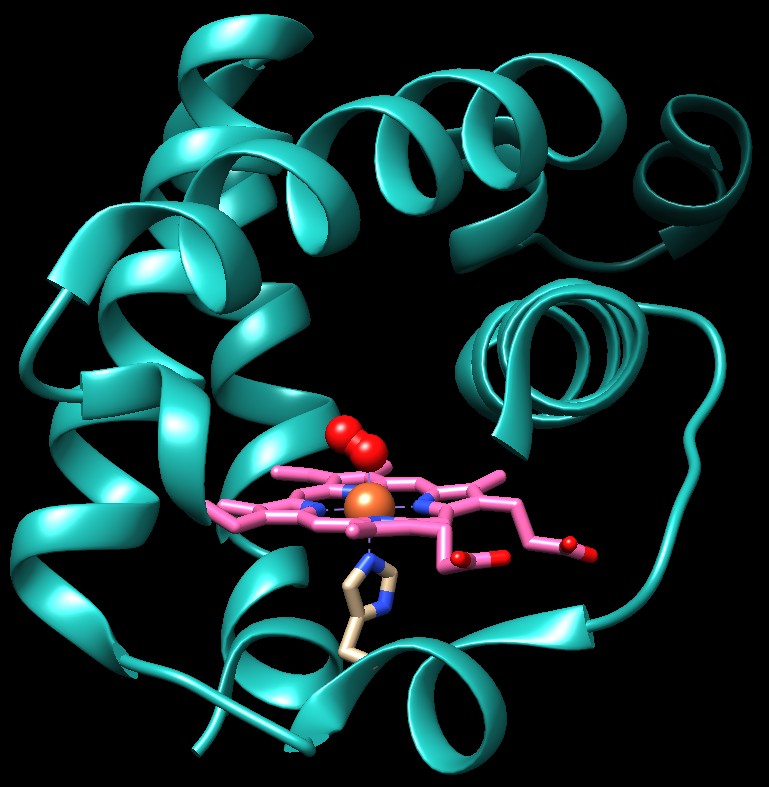
stata identificata una classe di cugini dell'emoglobina, le cosiddete
emoglobine troncate, come l'emoglobina mostrata qui sotto (filePDB
1idr) nella quale molte porzioni della
struttura classica sono state eliminate. L'unica caratteristica che
è sempre conservata in questo sottogruppo di proteine è
l'amminoacido di istidina che si lega al ferro dell'eme da sotto.
. . . . . . . . . . . . . . 
 L'unione
fa la forza L'unione
fa la forza
L'emoglobina è una straordinaria macchina molecolare che usa
il movimento di piccole variazioni strutturali per regolare la sua azione.
L'ossigeno O2
non si lega contemporaneamente ai quattro gruppi eme nell'emoglobina.
Il primo gruppo eme che lega l'ossigeno produce delle piccole modifiche
nella struttura 3D della catena proteica a cui è legato. Questi
cambiamenti strutturali premono leggermente le catene vicine che assumono
una forma diversa, e le inducono a legare l'ossigeno più facilmente.
Così, è difficile legare la prima molecola di ossigeno,
ma legare la seconda, la terza e la quarta molecola di ossigeno diventa
via via sempre più facile. Questo aiuta molto la funzione dell'emoglobina.
Quando il sangue è nei polmoni, dove l'ossigeno è abbondante,
l'ossigeno si lega facilmente alla prima subunità e poi rapidamente
riempe tutte le subunità rimanenti. Poi, quando il sangue circola
attraverso il corpo, il livello di ossigeno scende velocemente mentre
aumenta quello di biossido di carbonio. In questo ambiente, l'emoglobina
rilascia l'ossigeno legato. Appena viene rilasciata la prima molecola
di ossigeno, la proteina comincia a cambiare la sua forma. Questo induce
i tre ossigeni rimanenti ad essere rilasciati rapidamente. In questo
modo, l'emoglobina raccoglie il carico di ossigeno più grande
possibile nei polmoni, e lo consegna tutto dove e quando ce n'è
bisogno.
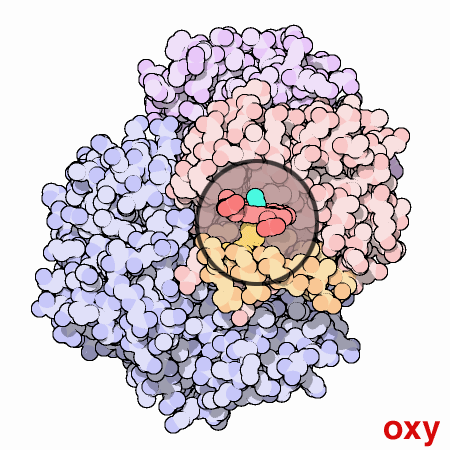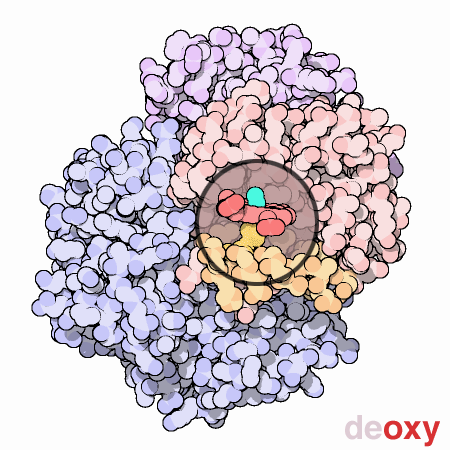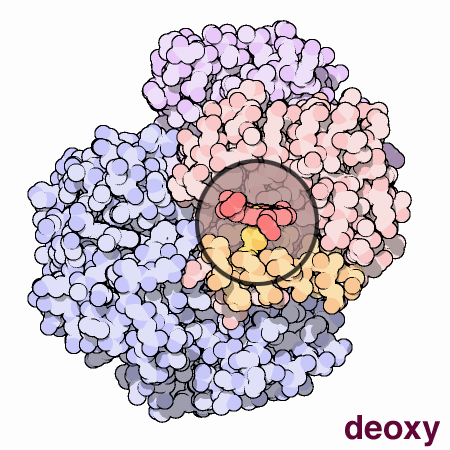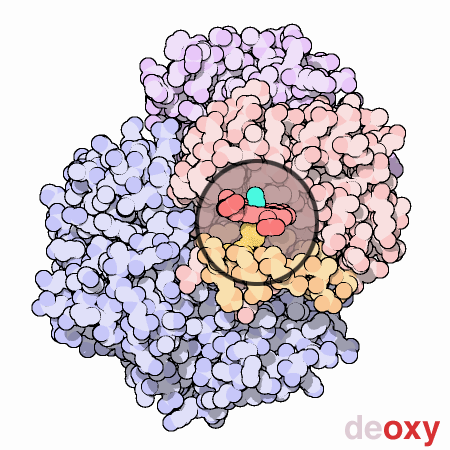
Nella figura animata qui a lato, il gruppo eme di una subunità,
mostrato nella piccola finestra circolare, viene mantenuto fermo in
modo che si possa vedere come la proteina gli si muove attorno quando
l'ossigeno si lega. La molecola di ossigeno è mostrata in azzurro.
Quando l'ossigeno si lega all'atomo di ferro nel centro dell'eme, tira
verso l'alto insieme al ferro anche l'amminoacido istidina che si trova
sul lato inferiore dell'eme. Questo sposta la posizione di un'intera
alfa-elica, mostrata qui in arancione sotto l'eme. Questo moto si propaga
a tutta la catena proteica e alle altre catene, provocando alla fine
il grande moto di oscillazione delle due subunità mostrate in
blu. Le due strutture mostrate qui sono prese dai file PDB 2hhb
e 1hho.
 Emoglobine difettose
Emoglobine difettose
I geni che codificano per le catene proteiche dell'emoglobina
mostrano piccole differenze all'interno delle diverse popolazioni umane,
così la sequenza degli amminoacidi dell'emoglobina è lievemente
diversa da persona a persona. Nella maggior parte dei casi queste variazioni
non incidono sulla funzione della proteina e spesso non si notano nemmeno.
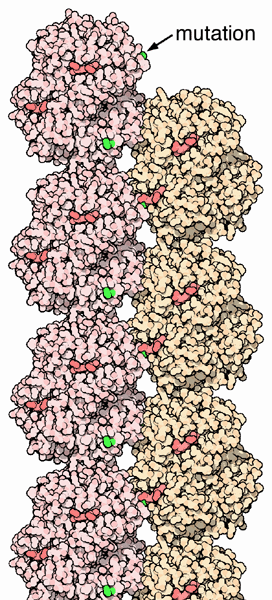
In alcuni casi, però, gli amminoacidi mutati producono grandi
variazioni strutturali. Un tipico esempio è quello dell'anemia
falciforme, dove l'acido glutammico 6 (codificato dalla tripletta
GAA) nella catena beta è sostituito da una valina (codificata
dalla tripletta GUA quindi una mutazione di un solo nucleotide). Questo
mutazione induce le molecole di emoglobina deossigenata ad ancorarsi
l'una all'altra, come si può vedere nella figura qui a fianco
(file PDB 2hbs). Si creano così
fibre rigide di emoglobina all'interno dei globuli rossi che si deformano
e, invece della normale forma a disco liscio, assumono una forma a C
o a falce. Le cellule deformate sono fragili e spesso si rompono
portando ad una perdita di emoglobina nota come anemia falciforme. Questa
condizione può sembrare che debba essere sempre un grave svantaggio,
ma in una circostanza almeno, diventa un vantaggio. I parassiti che
provocano la malattia tropicale della malaria, passano parte del loro
ciclo vitale nei globuli rossi, ma non possono vivere nei globuli rossi
falciformi pieni di fibre. Così le persone con l'anemia falciforme
sono resistenti alla malaria.
Altre circostanze che portano ad emoglobine difettose derivano da un
errore nella produzione delle proteine alfa o beta. La struttura dell'emoglobina
richiede un'uguale produzione di entrambe le proteine. Se una di queste
proteine manca, si produce una condizione chiamata talassemia.
Esplorando la struttura
Qui sotto si può vedere il sito di legame
dell'ossigeno in due strutture di emoglobina umana.
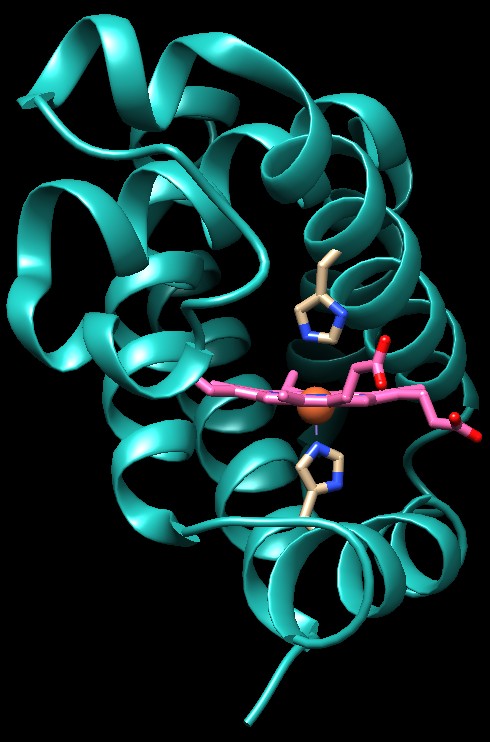
La struttura sulla sinistra (file PDB 2hhb)
mostra l'emoglobina senza ossigeno legato. Il gruppo eme è visto
di lato con l'atomo di ferro arancione. Si può vedere che c'è
un amminoacido di istidina che si trova molto vicino al lato inferiore
dell'eme, e forma un legame con l'atomo di ferro. Si noti che l'atomo
di ferro, a causa di questo legame, si trova leggermente sotto il piano
dell'eme.
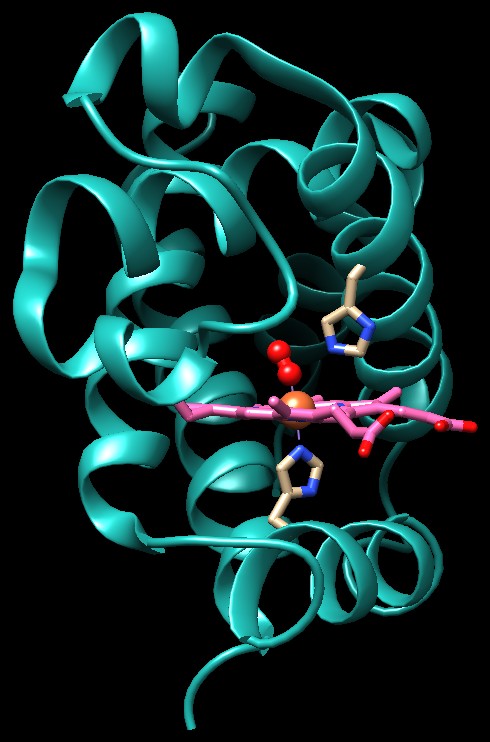
La struttura sulla destra (file PDB 1hho)
mostra che, quando l'ossigeno si lega al ferro, lo tira verso l'alto
portandolo leggermente sopra il piano del'eme. Il ferro, a sua volta,
tira verso l'alto l'istidina, la quale trascina e deforma l'alfa elica
in basso.
Questi movimenti sono trasmessi alle altre subunità e alla fine
provocano la grande variazione strutturale che cambia la forza di legame
dei siti vicini. La deformazione fa avvicinare anche le istidine delle
altre subunità al loro eme spingendo il ferro verso l'alto nel
piano dell'eme e aumentando la sua capacità di legare l'ossigeno.
Notate che l'archivio PDB 1hho contiene solamente due delle quattro
catene dell'emoglobina. Per vedere il tetramero completo, scaricate
il file PDB 1hho4.
 Bibliografia Bibliografia
Perutz, M.F. (1978): Hemoglobin Structure
and Respiratory Transport. Scientific American, 239 (6).
Squires, J.E. (2002): Artificial Blood. Science 295,
p.1002.
Vichinsky, E. (2002): New therapies in sickle cell disease. Lancet
24, p. 629.
|
|
|
