|
|
Amminoacil-tRNA sintetasi |
|||||
|
Molecola del mese di aprile 2001 Gli enzimi amminoacil-tRNA sintetasi garantiscono che solo gli amminoacidi corretti vengano usati per la sintesi delle proteine 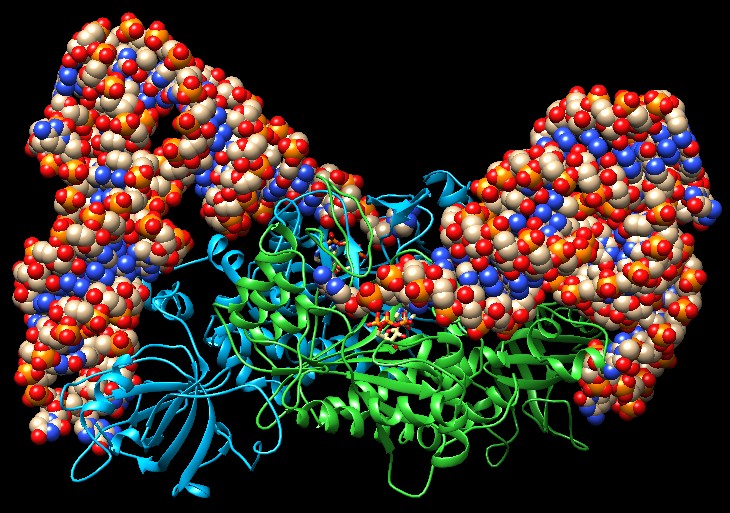 Introduzione IntroduzioneQuando un ribosoma fa appaiare l'anticodone CGC del tRNA con il codone GCG dell'mRNA, si aspetta che il tRNA trasporti alanina alla sintesi di proteine. Del resto non ha nessun modo di verificare se questo avviene effettivamente. Infatti il tRNA (mdm 3/2001) si lega al proprio amminoacido molto prima di giungere al ribosoma. Il giusto appaiamento è realizzato da una serie di enzimi chiamati amminoacil-tRNA sintetasi. Questi enzimi legano ad ogni tRNA il giusto amminoacido e così consentono ad ogni tRNA di tradurre in modo corretto il codice genetico di nucleotidi del DNA nel codice di amminoacidi delle proteine. Venti enzimi diversi La maggior parte delle cellule sintetizzano venti diversi amminoacil-tRNA sintetasi, uno per ogni amminoacido. Questi venti enzimi sono molto diversi tra loro dato che sono ottimizzati per funzionare con il loro amminoacido e per quel set di tRNA che sono deputati a trasportarlo. L'enzima mostrato qui sopra, che installa acido aspartico sul suo tRNA (file PDB 1asz), è un dimero composto di due subunità identiche (azzurra e verde), che legano due molecole di tRNA (sfere colorate). Altri enzimi sono piccoli monomeri, altri ancora grossi monomeri o dimeri o tetrameri formati da subunità uguali o diverse. Alcuni hanno una forma bizzarra come l'enzima della serina (file PDB 4rqe), mostrato qui sotto, che possiede su ognuno dei due monomeri un lungo braccio per riconoscere anche altri punti del tRNA. Notate che nel sito attivo dei due monomeri è presente ATP e l'amminoacido serina (Ser) che deve reagire prima con ATP e poi con il ribosio terminale del tRNA. . . . . 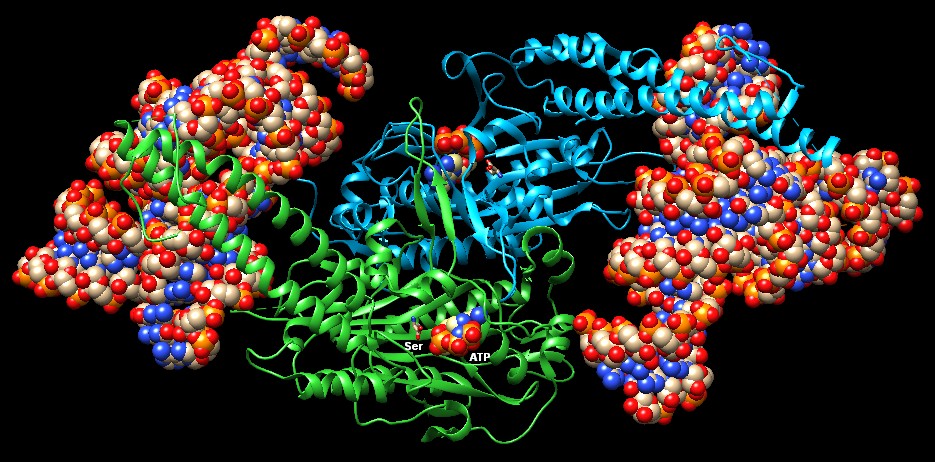
Trovare il proprio compagno Come è logico aspettarsi, molti di questi enzimi riconoscono il proprio tRNA dal suo anticodone, ma non sempre questo è possibile. Con la serina, per esempio, che è codificata da sei diversi codoni, è necessario un seril-tRNA sintetasi (mostrato qui sopra) che riconosca sei diversi tRNA con sei diversi anticodoni tra i quali AGA e GCU che sono del tutto diversi tra loro e inoltre che sappia riconoscere il tRNA della selenocisteina. Per questo le molecole di tRNA sono anche riconosciute in base ad alcuni tratti all'altro capo della molecola, sul terminale accettore, dove si lega l'amminoacido, o anche a porzioni diverse. In particolare la base 73 della sequenza sembra avere un ruolo cruciale in molti casi ed è stata chiamata base discriminante. In altri casi, d'altra parte, è del tutto ignorata. Notate anche che ogni enzima deve riconoscere le proprie molecole di tRNA e, nello stesso tempo, non si deve legare a nessuno degli altri tRNA. Così, ogni tRNA ha un set di interazioni positive che gli consentono di legarsi al giusto enzima e un set di interazioni negative che gli impediscono di legarsi ad ogni altro enzima. Per esempio, aspartil-tRNA sintetasi mostrato qui sopra (file PDB 1asz) riconosce la base discriminante e quattro basi vicino all'anticodone. Però, un'altra base, guanina 37, che non è usata per il legame, deve essere metilata per garantire che il tRNA non si leghi all'enzima arginil-tRNA sintetasi. Sorprese dall'analisi del genoma Recenti studi del genoma hanno rivelato una cosa sorprendente: alcuni organismi non hanno geni per tutti i venti enzimi amminoacil-tRNA sintetasi anche se usano tutti i venti amminoacidi per le loro proteine. La soluzione di questo paradosso ha rivelato che in realtà sono in atto meccanismi più complessi. Per esempio, alcuni batteri non hanno l'enzima che lega la glutammina al suo tRNA, ma un singolo enzima aggiunge acido glutammico sia al tRNA dell'acido glutammico che a quello della glutammina. Poi un secondo enzima converte l'acido glutammico in glutammina sul tRNA che deve avere glutammina, ripristinando il corretto appaiamento. Approcci diversi per lo stesso problema Nella figura qui sotto sono illustrati cinque complessi tra amminoacil-tRNA sintetasi e il rispettivo tRNA disposti in modo che il tRNA (rosso) abbia sempre lo stesso orientamento. Ogni enzima lega il suo tRNA con una diversa angolazione. I primi tre enzimi di isoleucina (file PDB 1ffy), valina (file PDB 1gax) e glutammina (file PDB 1euq) abbracciano il tRNA prendendolo per l'anticodone (in basso) e portando l'altra estremità del tRNA (in alto a destra) che deve legare l'amminoacido, nel sito attivo dell'enzima. Questi enzimi adottano una logica simile per la reazione e sono detti di tipo 1: abbracciano il tRNA in modo simile e aggiungono l'amminoacido all'OH 2' del ribosio terminale del tRNA. Gli ultimi due enzimi di fenilalanina (file PDB 1eiy) e treonina (file PDB 1qf6) e quello dell'acido aspartico mostrato all'inizio (file PDB 1asz) appartengono ad una seconda classe di enzimi, detti di tipo 2. Questi abbracciano il tRNA dal lato opposto e aggiungono l'amminoacido all'OH 3' del ribosio terminale del tRNA. In realtà l'enzima della fenilalanina ha caratteristiche intermedie tra quelli di tipo 1 e 2 perché abbraccia il tRNA dal lato opposto (tipo 2), ma aggiunge l'amminoacido all'OH 2' (tipo 1). 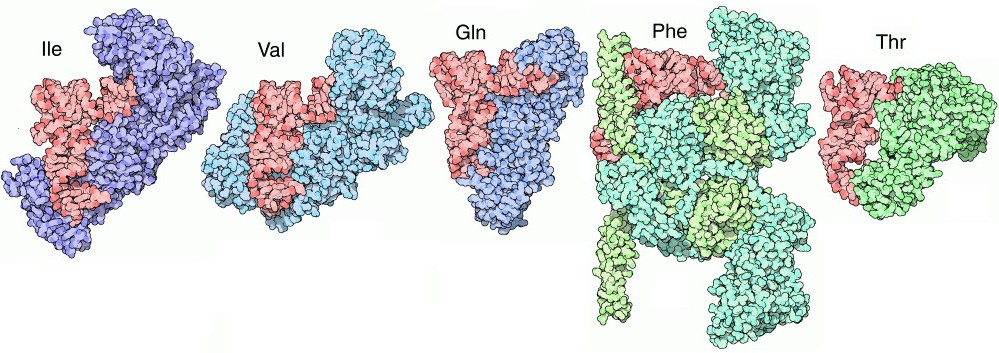 Alta fedeltà Gli enzimi amminoacil-tRNA sintetasi devono operare con la massima accuratezza dato che, ogni volta che fanno un errore, provocano la sintesi di una proteina con un amminoacido errato. Mediamente fanno un errore ogni 10000 volte, ma, visto che la maggior parte degli amminoacidi sono molto diversi uno dall'altro, questo livello di accuratezza non è molto difficile da ottenere. Alcuni amminoacidi, però, sono molto simili tra loro e così gli enzimi devono escogitare tecniche speciali per legare l'amminoacido giusto. La coppia di amminoacidi isoleucina (ile) e valina (val), per esempio costituisce una sfida notevole per questi enzimi. . . . . . . 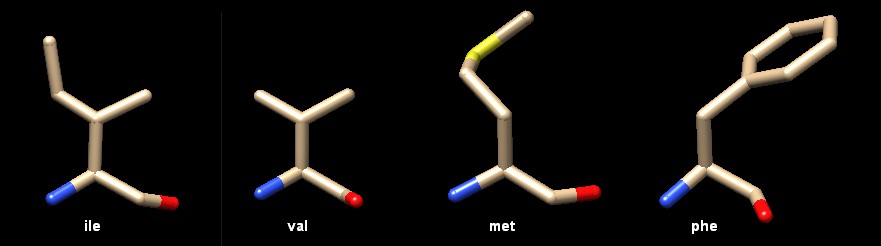
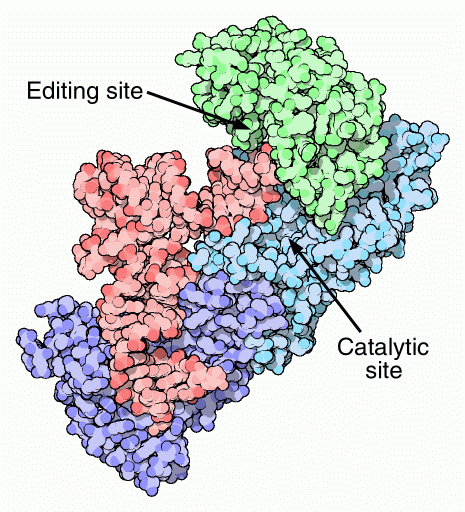 L'isoleucina
è un amminoacido apolare riconosciuto da una tasca di forma opportuna
nell'enzima. Questa è troppo piccola per accogliere altri amminoacidi
apolari più grandi come metionina (met) o fenilalanina
(phe) e troppo idrofobica per accogliere amminoacidi polari. L'isoleucina
è un amminoacido apolare riconosciuto da una tasca di forma opportuna
nell'enzima. Questa è troppo piccola per accogliere altri amminoacidi
apolari più grandi come metionina (met) o fenilalanina
(phe) e troppo idrofobica per accogliere amminoacidi polari. La valina, però, ha una catena apolare appena un po' più piccola dell'isoleucina, con un solo gruppo metilico in meno, ed entra nella tasca dedicata alla isoleucina e così l'enzima aggiunge valina una volta su 150. Questi errori sono decisamente troppi e quindi è necessario intervenire per correggerli. L'enzima isoleucil-tRNA sintetasi mostrato qui a lato (file PDB 1ffy ) risolve questo problema con un secondo sito attivo di correzione (editing) che può staccare l'amminoacido. L'isoleucina, un po' più grande, non può entrare in questo secondo sito, ma la valina sì. L'amminoacido errato, così, viene staccato lasciando il tRNA pronto per legare quello corretto, isoleucina. Questo passaggio di correzione migliora l'azione dell'enzima che così compie un errore ogni 3000 volte. Esplorando la struttura Questi enzimi non trattano con gentilezza il tRNA. Un esempio interessante di questa rudezza si ha con il glutaminil-tRNA sintetasi mostrato qui sotto (file PDB 1gtr). L'enzima stringe con forza l'anticodone del tRNA in basso facendo allargare le tre basi azotate (evidenziate in giallo, verde e magenta) per poterle riconoscere con tre diverse tasche. Sul terminale opposto, l'enzima spaia una base all'inizio della catena (che sporge verso l'alto) e costringe il terminale accettore (a destra) a formare una piega molto stretta a forcina con i nucleotidi viola, arancione, verde e giallo (terminale) in sequenza. In questo modo l'OH 2' (rosso) del ribosio terminale (giallo) entra nel sito attivo dove si trovano l'amminoacido (non mostrato) e l'ATP che devono reagire. . . . . . . .. . . . 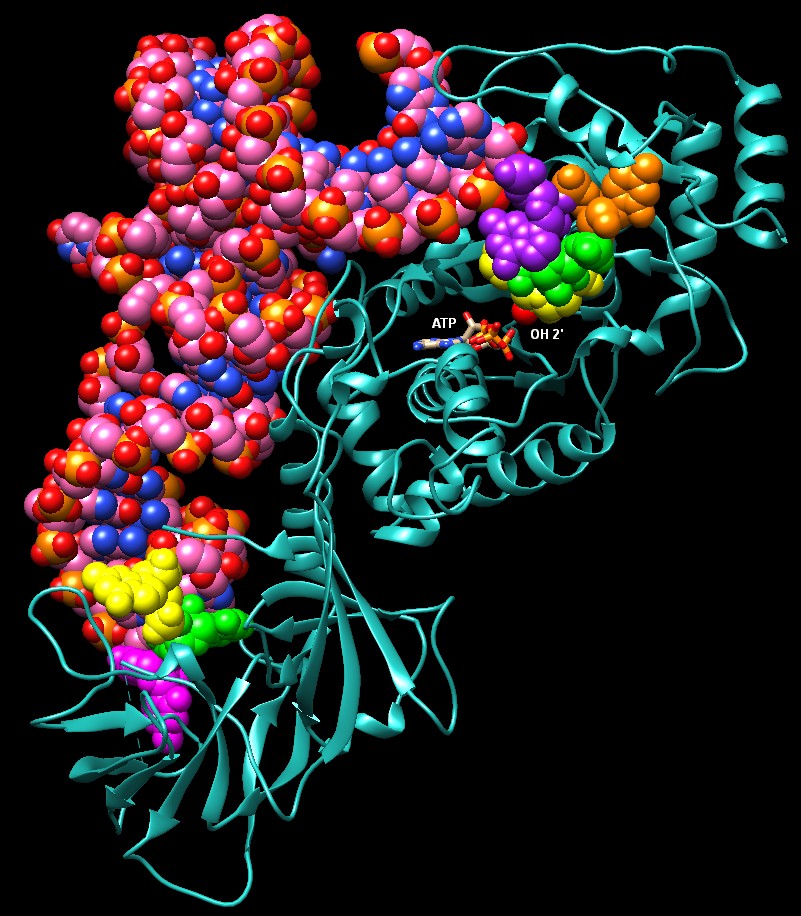 Michael Ibba and Dieter Soll (2000) Aminoacyl-tRNA Synthesis. Annual Review of Biochemistry 69, 617-650. John G. Arnez and Dino Moras (1997) Structural and Functional Considerations of the Aminoacylation Reaction. Trends in Biochemical Sciences 22, 211-216. Stephen Cusak (1995) Eleven Down and Nine to Go. Nature Structural Biology 2, 824-831. Jonathan J. Burbaum and Paul Schimmel (1991) Structural Relationships and the Classification of Aminoacyl-tRNA Synthetases. Journal of Biological Chemistry 266, 16965-16968.
|
||||||
|
|
||||||
